Cell Membrane and Transport
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
The fundamental requirement for any living system to exist is the establishment of a boundary. Without a physical barrier separating the chaos of the external universe from the highly ordered biochemistry of the internal environment, life diffuses into nothingness. The plasma membrane is this definitive border. It is not a static wall of plastic wrap, but a bustling, dynamic frontier that constantly negotiates what enters and exits the cell. As a future biology teacher, mastering the mechanics of this boundary allows you to explain to a student why a wilted plant snaps back to life after a rainstorm, why drinking seawater accelerates dehydration, and how their very neurons fire to process the lesson you are teaching.
To understand how the cell regulates its contents, we must first look at the materials of its construction. The plasma membrane is composed primarily of a phospholipid bilayer.
Why a bilayer? The answer lies in the chemical duality of the building blocks. Phospholipids are amphipathic molecules, meaning they harbor two contradictory chemical personalities within the same structure. A single phospholipid molecule contains a hydrophilic phosphate head, which aggressively seeks interaction with water, and two hydrophobic fatty acid tails, which violently repel it.
When you place thousands of these molecules into an aqueous environment, they self-assemble. The hydrophobic tails of membrane phospholipids face inward to form a water-excluding core, escaping the water entirely. Consequently, the hydrophilic heads of membrane phospholipids face outward toward the aqueous extracellular fluid and the aqueous cytoplasm.
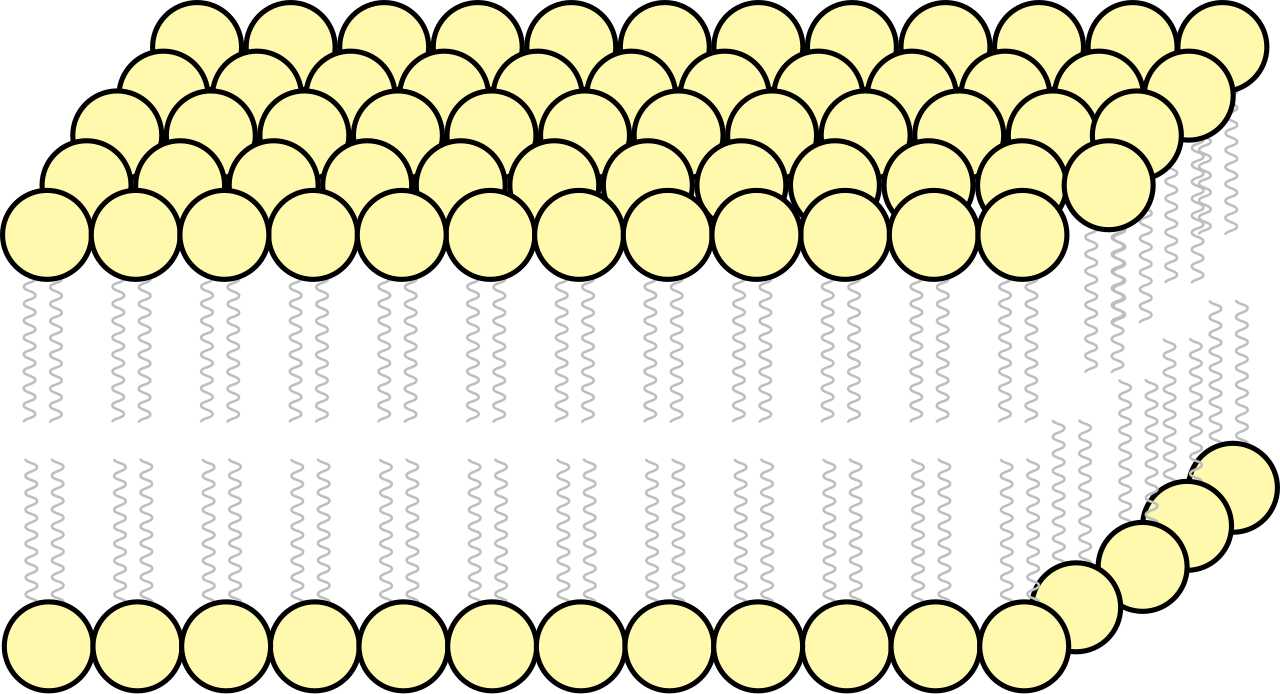
This structure is highly mobile. The fluid mosaic model describes the dynamic structural organization of the plasma membrane. The lipid molecules are not locked in place; phospholipids in the plasma membrane can continuously move laterally within their own monolayer, drifting like buoys on a molecular sea.
To maintain the perfect consistency of this lipid sea—neither too solid nor too fluid—nature employs chemical buffers. Cholesterol molecules are embedded within the hydrophobic core of the animal cell membrane. As a temperature buffer, cholesterol buffers membrane fluidity by preventing the tight packing of lipids at low environmental temperatures, keeping the membrane from freezing into a solid matrix. Conversely, cholesterol restricts the excessive lateral movement of phospholipids at high environmental temperatures, preventing the membrane from melting and dissolving.
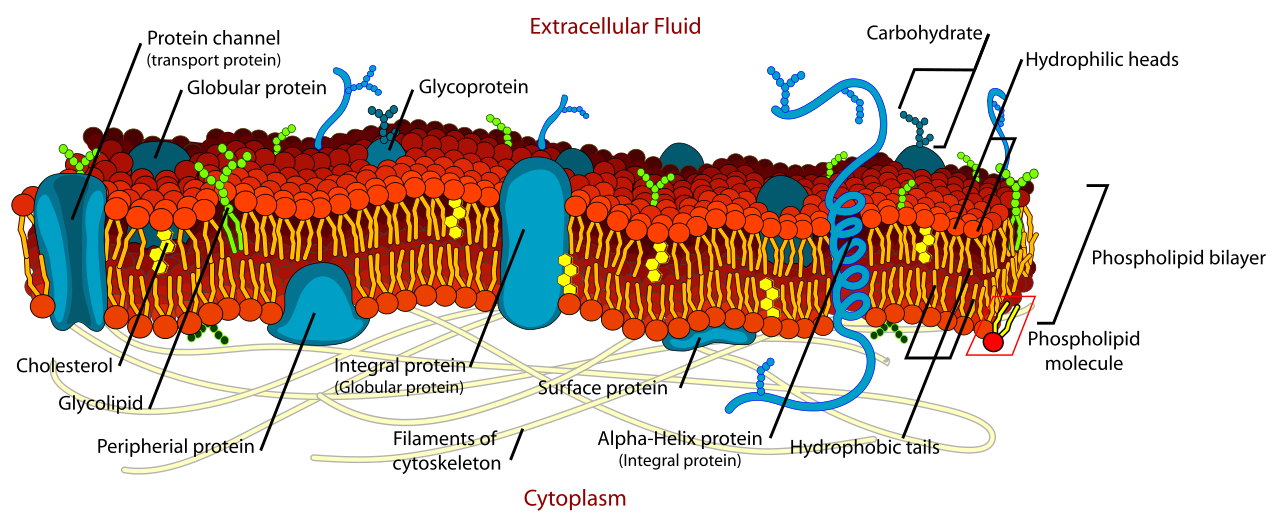
The Membrane Mosaic: Proteins and Carbohydrates
If phospholipids are the sea, proteins are the ships, anchors, and ports.
- Integral membrane proteins embed directly into the hydrophobic core of the phospholipid bilayer.
- Transmembrane proteins are a specific type of integral protein spanning the entire width of the plasma membrane, creating physical bridges between the inside and outside of the cell.
- Peripheral membrane proteins attach loosely to the exterior or interior surfaces of the plasma membrane, often serving as anchor points for the cytoskeleton or relay stations for intracellular signaling.
Extending from the exterior of this lipid-protein matrix is a molecular "forest" of sugar chains. Carbohydrate chains attached to membrane proteins form glycoproteins, while carbohydrate chains attached to membrane lipids form glycolipids. These sugar complexes are the molecular ID tags of the cell; membrane glycoproteins function in cellular identification and cell-to-cell recognition. This is how an immune cell knows to ignore a liver cell but attack a foreign bacterium.
Because of the thick, oily interior of the bilayer, the phospholipid bilayer exhibits selective permeability. The rules of crossing are dictated entirely by physics and chemistry.
Small nonpolar molecules diffuse directly across the hydrophobic core of the plasma membrane. Because they lack charge and are tiny, the hydrophobic tails do not repel them. For example, oxygen gas diffuses directly through the phospholipid bilayer without the aid of membrane proteins. This is biologically vital: your cells do not have to wait for a protein "door" to open to receive the oxygen they need for cellular respiration.
In stark contrast, charged ions cannot readily cross the hydrophobic core of the plasma membrane. Their electrical charge makes them entirely incompatible with the nonpolar lipid tails. Furthermore, large polar molecules require transport proteins to cross the plasma membrane, as their size and polarity prevent them from slipping through the lipid matrix.
The Mechanics of Passive Transport
When molecules move down a gradient without energy, we call it passive transport.
Passive transport is the movement of substances across a membrane without the expenditure of cellular metabolic energy. By the laws of thermodynamics, passive transport forces molecular movement exclusively down a concentration gradient (from an area of high concentration to an area of low concentration).
There are two primary modes of passive transport:
- Simple diffusion is a form of passive transport lacking the involvement of transport proteins. (e.g., Oxygen slipping through the lipids).
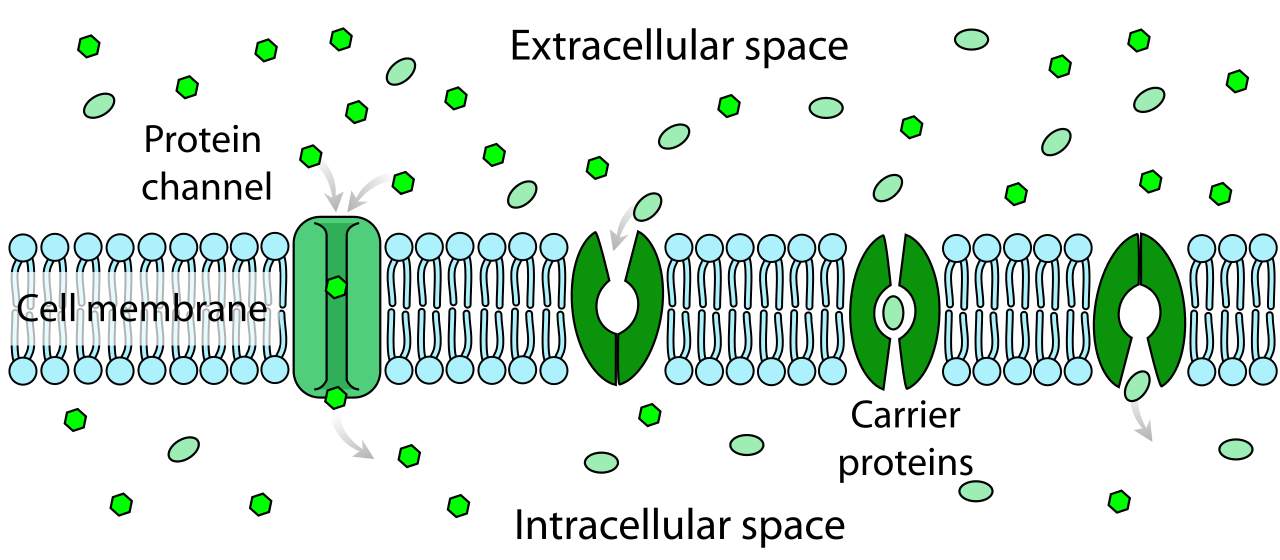
- Facilitated diffusion is a form of passive transport requiring specific transmembrane proteins. This is for the ions and large polar molecules that want to cross but need a bypass through the hydrophobic core.
Within facilitated diffusion, the transmembrane proteins take two main forms:
- Channel proteins provide a hydrophilic corridor for specific ions or molecules to traverse the membrane. Think of them as molecular tunnels.
- Carrier proteins undergo a conformational shape change to move specific molecules across the plasma membrane. Think of them as revolving doors that physically grab a molecule, shift their 3D shape, and dump the molecule on the other side.
Life cannot survive on passive equilibrium alone. Sometimes, a cell must hoard nutrients or expel toxins against their natural gradient.
Active transport requires the direct or indirect expenditure of cellular metabolic energy, and crucially, active transport drives molecular movement against a concentration gradient (from low to high concentration).
Primary Active Transport
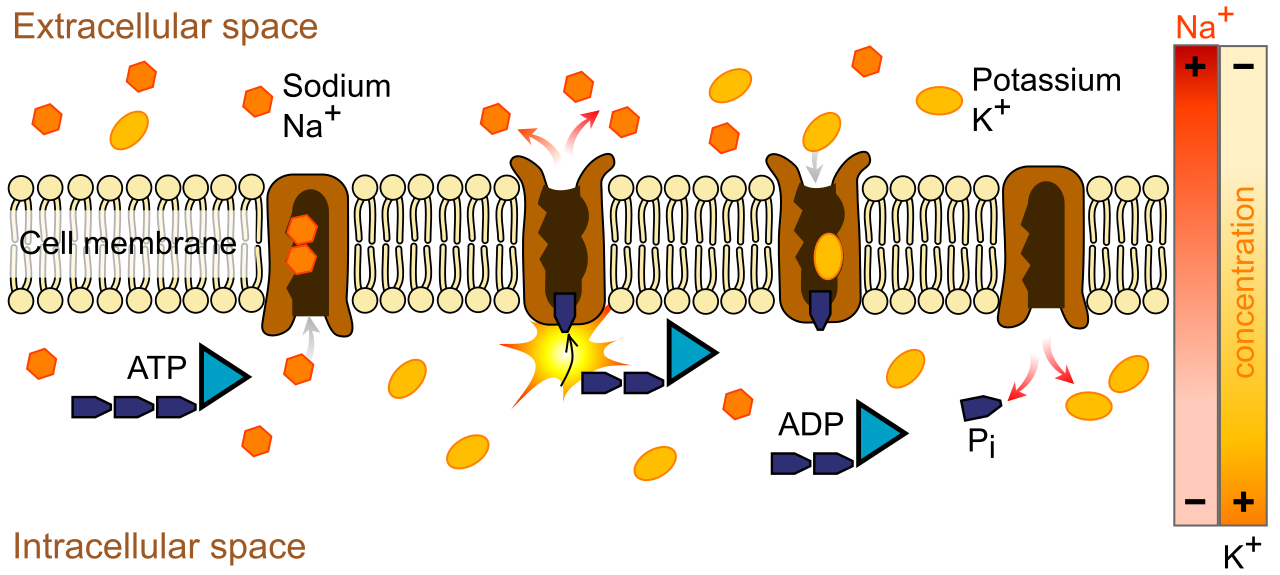
Primary active transport utilizes the hydrolysis of ATP to directly power the movement of ions across a membrane.
The textbook, canonical example of this is the nerve cell's battery charger. The sodium-potassium pump is an example of a primary active transport protein. To maintain the electrochemical gradient necessary for nervous system function, this pump operates with rigid, mechanical precision: the sodium-potassium pump exports exactly three sodium ions out of the cell during one operational cycle, while simultaneously, the sodium-potassium pump imports exactly two potassium ions into the cell during one operational cycle.
Secondary Active Transport
Once a primary pump has established a massive concentration gradient, the cell can use that stored potential energy to do work, much like water held behind a dam. Secondary active transport uses an established electrochemical gradient to power the transport of a different target molecule.
This relies on coupled transport proteins:
- Symporters are membrane transport proteins moving two different molecular species in the same physical direction. (e.g., sodium rushing back into the cell, dragging glucose along with it).
- Antiporters are membrane transport proteins moving two different molecular species in opposite physical directions.
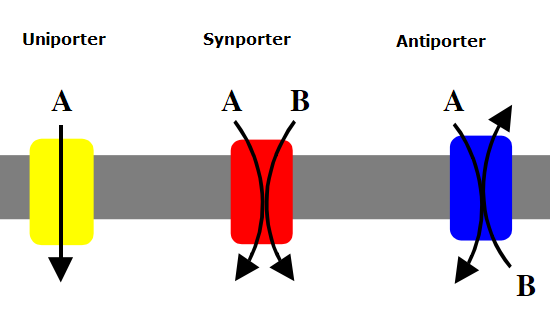
For biology students, water movement is often a hurdle. You must teach them to view water as a molecule that obeys the same rules of diffusion as everything else.
Osmosis is the facilitated diffusion of water molecules across a selectively permeable membrane. While a tiny amount of water can slip through the bilayer, bulk water movement requires doors. Aquaporins are specialized channel proteins facilitating the rapid transport of water across cell membranes.
To predict where water will go, we look at osmolarity. Osmolarity defines the total concentration of dissolved solute particles in a given volume of solution. The golden rule is simple: water moves osmotically from a region of lower osmolarity toward a region of higher osmolarity. Water always chases the solutes.
Tonicity: The Cellular Environment
We classify external environments based on their total solute concentration relative to the cell's interior.
| Environment | Definition | Cellular Outcome |
|---|---|---|
| Hypertonic | A hypertonic solution possesses a higher total solute concentration than the cytoplasm of a referenced cell. | A cell submerged in a hypertonic solution experiences a net loss of water. In animals, the cell shrivels. In plants, plasmolysis occurs when a plant cell membrane detaches from the cell wall due to water loss in a hypertonic environment. |
| Hypotonic | A hypotonic solution possesses a lower total solute concentration than the cytoplasm of a referenced cell. | An animal cell submerged in a hypotonic solution experiences a net gain of water, swelling until an animal cell submerged in a hypotonic solution can undergo cytolysis (bursting). Conversely, because of their rigid bounds, a plant cell submerged in a hypotonic solution develops turgor pressure against the rigid cell wall, keeping the plant upright. |
| Isotonic | An isotonic solution possesses an identical total solute concentration to the cytoplasm of a referenced cell. | A cell submerged in an isotonic solution experiences no net osmotic movement of water. Water enters and exits at equal rates. |
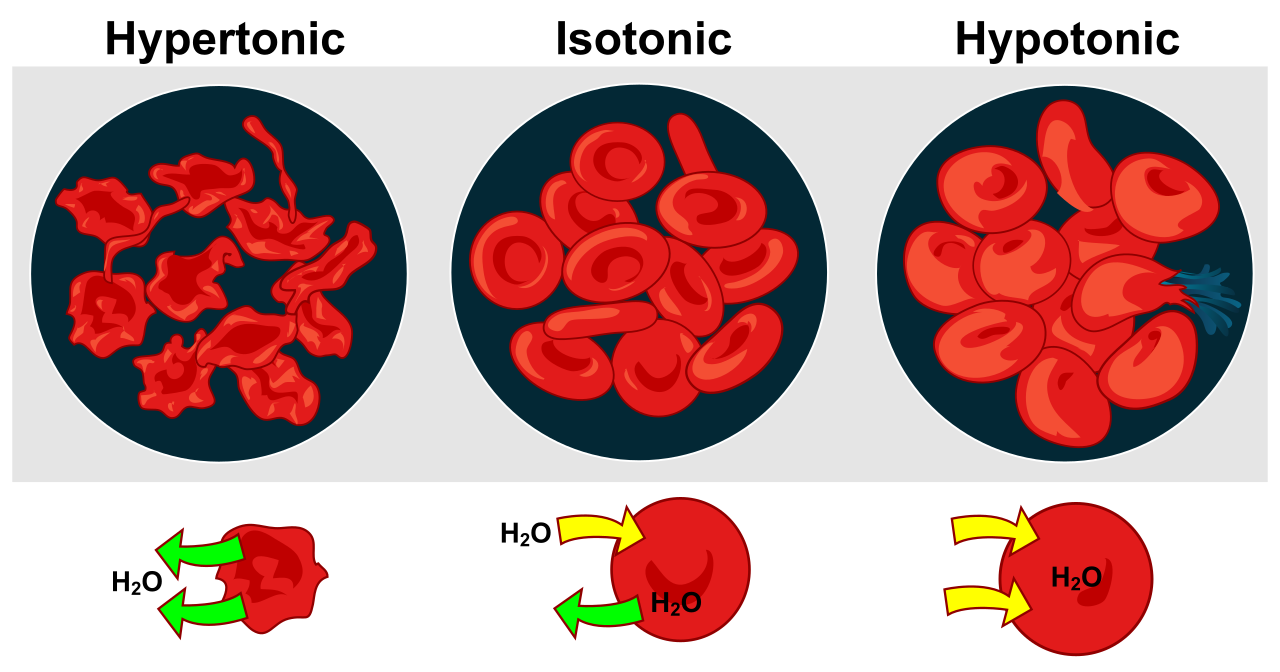
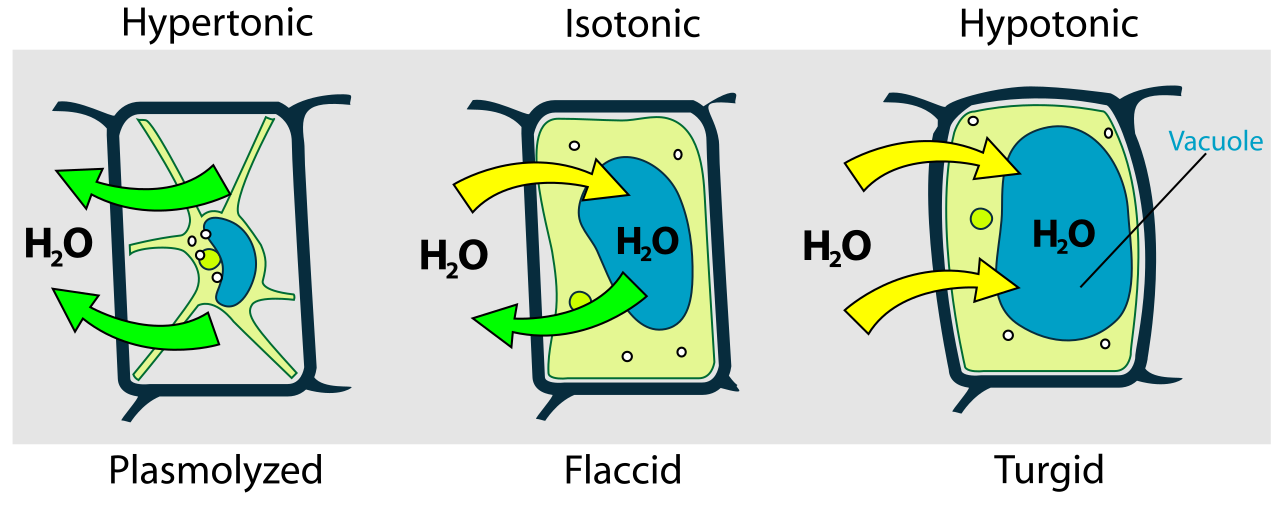
The Physics of Water Movement: Water Potential
To truly quantify this for plant physiology, we use a concept from physics. Water potential determines the physical direction of water movement across a biological membrane. The thermodynamic law dictates that water physically moves from a region of higher water potential toward a region of lower water potential.
Total water potential (Ψ) is the mathematical sum of the solute potential (ΨS) and the pressure potential (ΨP) within a system. Ψ=ΨS+ΨP
Here is how you break this down for your future students:
- Pure water in an open container possesses a solute potential of exactly zero. This is the maximum possible baseline.
- The addition of dissolved solutes structurally decreases the solute potential of a solution. Because the baseline is zero, the solute potential of any aqueous solution is always a negative mathematical value. The more stuff you dissolve in water, the more negative the solute potential becomes, and the stronger it "pulls" water toward it.
- Pressure potential measures the physical pressure exerted on a liquid solution. In an open beaker, this is zero. In a plant cell, the rigid wall pushes back against the swelling membrane, creating a positive pressure potential that eventually stops the inward flow of water, regardless of solutes.
Sometimes a cell needs to move entire macromolecules, complex fluids, or even whole bacteria. Proteins and channels are too small for this task. Bulk transport mechanisms move massive molecules or large volumes of material across the plasma membrane. Because of the massive rearrangement of the membrane and cytoskeleton required, all forms of bulk transport require the expenditure of cellular metabolic energy.
Exiting the Cell
Exocytosis is the cellular process of exporting materials via the fusion of internal vesicles with the plasma membrane. When a nerve cell releases neurotransmitters, or a pancreas cell secretes insulin, vesicles migrate from the Golgi apparatus, meld their phospholipid bilayers with the plasma membrane, and spill their contents into the extracellular space.
Entering the Cell
Endocytosis is the cellular process of internalizing materials through the inward folding of the plasma membrane. The membrane pinches inward to form a new internal vesicle. There are three distinct variations:
- Phagocytosis is a distinct form of endocytosis utilized by cells to engulf large solid particles or whole microscopic organisms. This is "cellular eating," employed by white blood cells to destroy invading pathogens.
- Pinocytosis is a distinct form of endocytosis utilized by cells to internalize extracellular fluid and dissolved solutes. This is "cellular drinking," a way for the cell to constantly sample the surrounding chemical environment.
- Receptor-mediated endocytosis initiates exclusively when specific target molecules bind to surface receptor proteins. This is highly specific. When a cell needs to pull in massive amounts of cholesterol from the bloodstream, it relies on these receptors. Once bound, the binding of specific ligands to surface receptors triggers the formation of a clathrin-coated intracellular vesicle, effectively boxing up the target molecules in a dedicated molecular shipping container and pulling them safely inside.
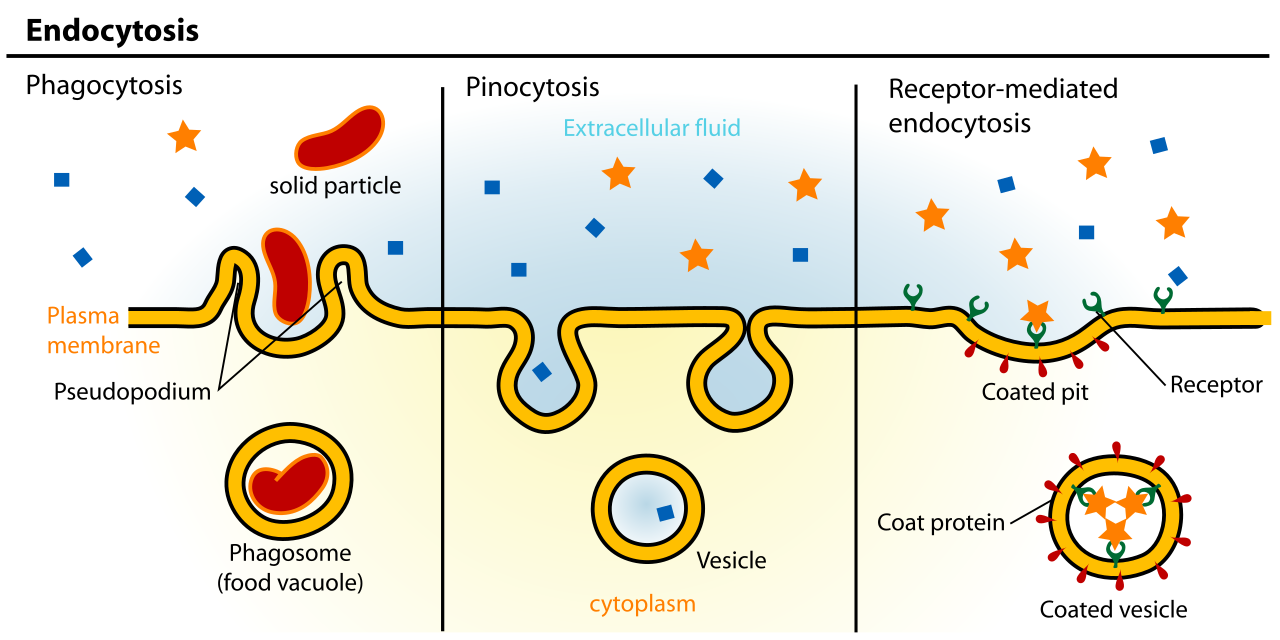
By understanding these mechanisms—from the tiny, silent slipping of an oxygen molecule across the lipid bilayer, to the violent, energy-intensive engulfing of a bacterium by a macrophage—you equip your future students with the blueprints to cellular life itself.