Cell Communication
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
A human body is not a single living entity; it is a meticulously coordinated collective of over thirty trillion individual cells. Without wires, without a central processor physically touching each unit, these microscopic biological machines must breathe, eat, divide, and defend themselves in absolute synchrony. The mechanism that prevents this vast collective from collapsing into biological anarchy is cell communication—a physical, chemical reality where molecules float across fluid expanses to act as keys in highly specific locks.
As an aspiring biology teacher, you will soon stand in front of a classroom and face the challenge of explaining how cells "talk." High school students often imagine cell signaling as magical radio waves. Your job is to ground this process in physical, tangible mechanics: shapes shifting, chemical tags attaching, and molecular cascades amplifying. The Praxis (5236): Biology: Content Knowledge exam demands a rigorous understanding of these specific mechanics. We are going to explore the canonical architecture of cellular communication, breaking it down into the exact vocabulary, mechanisms, and pathways you must master.
When you manage a classroom, you use different types of communication depending on the distance and the target. You might make an announcement to the whole school over the PA system, whisper to a student in the front row, or leave a sticky note on your own desk as a reminder. Cells operate on the exact same logic.
At the center of all these pathways is the ligand.
A ligand is a molecule that specifically binds to another target molecule. It is the molecular "word" being spoken.
Depending on how far that word needs to travel, biological systems utilize different strategies:
- Autocrine signaling targets the cell that secretes the signal. Think of this as a cell leaving a sticky note for itself. It releases a ligand that binds to its own surface receptors, often to self-regulate its own biological activity.
- Paracrine signaling targets cells in the immediate vicinity of the emitting cell. This is the cellular equivalent of talking to your neighbors. The ligands diffuse over short distances through the extracellular matrix.
- Synaptic signaling is a specialized form of paracrine signaling between a neuron and a target cell. When an electrical impulse reaches the end of a neuron, it triggers the release of specialized ligands. Neurotransmitters are the signaling molecules used in synaptic signaling. They cross the tiny synaptic gap to bind to the neighboring cell, ensuring lightning-fast communication in the nervous system.
- Endocrine signaling uses the circulatory system to transport signaling molecules to distant target cells. This is the PA system of the body. Hormones are the specific signaling molecules used in endocrine signaling. Because they travel through the bloodstream, they can act on multiple diverse organ systems simultaneously, though they take longer to reach their destinations than paracrine signals.
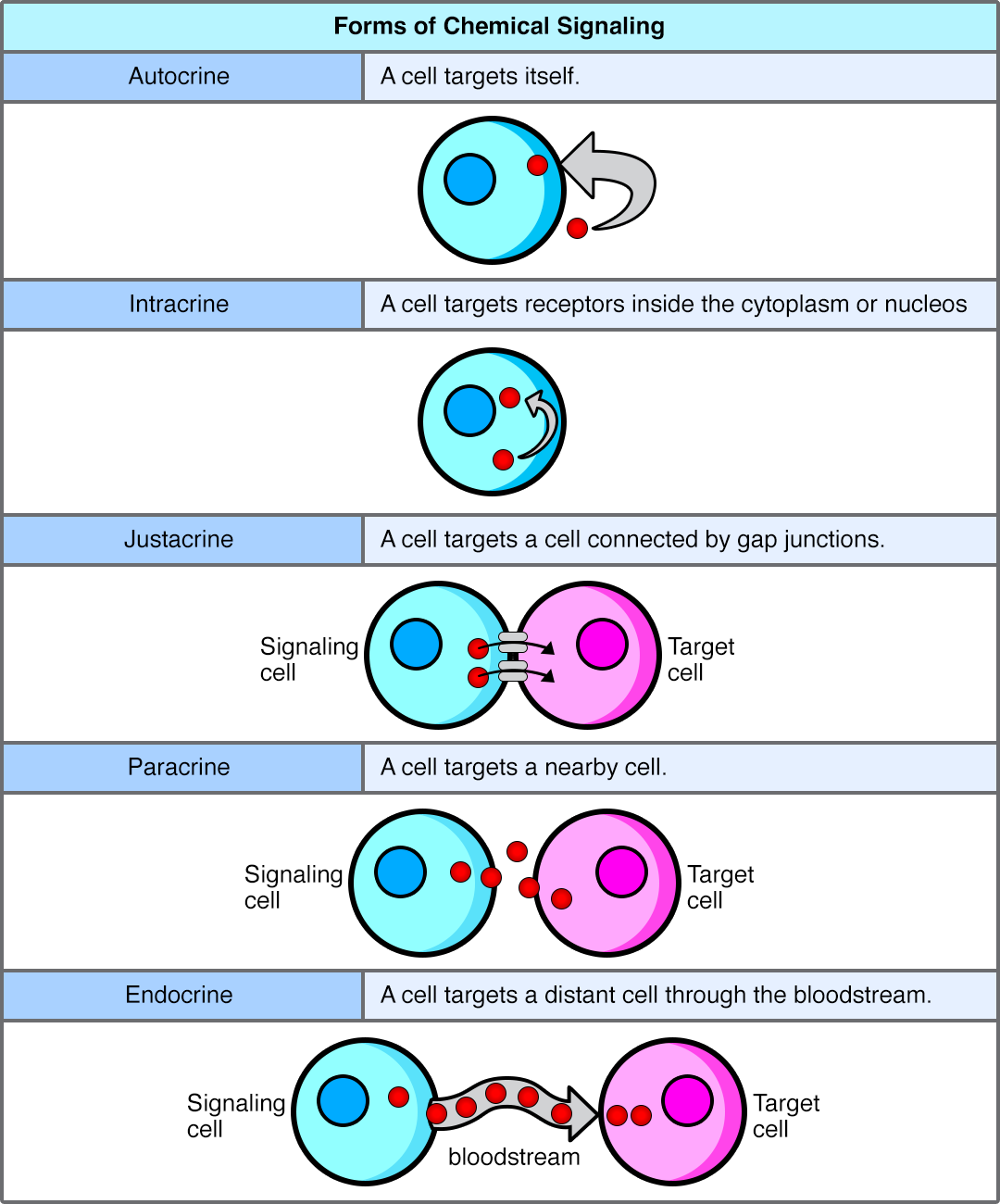
- Juxtacrine signaling requires direct physical contact between interacting cells. The cells must touch. Sometimes, this happens through structural portals that physically link the cytoplasm of two adjacent cells.
- Gap junctions in animal cells allow small signaling molecules to pass directly between adjacent cells.
- Plasmodesmata in plant cells allow small signaling molecules to pass directly between adjacent cells. (Remember: plant cells have rigid cell walls, so these plasmodesmata act as vital tunnels through that tough barrier).
In 1971, Earl W. Sutherland won the Nobel Prize for proving that cell communication isn't just one event—it is a sequence. The three primary stages of cell signaling are reception, transduction, and response.
Let's walk through the physical reality of each stage.
Stage 1: Reception – The Molecular Handshake
Before a cell can respond to a signal, it has to "hear" it. A receptor is a protein molecule that receives and binds to a specific signaling molecule.
Because a receptor is a protein, it has a highly specific three-dimensional shape. When the correct ligand bumps into it with the right orientation and energy, they bind. The immediate, physical consequence of this binding is paramount: Ligand binding causes a conformational change in the receptor protein.
A conformational change is an alteration in the three-dimensional shape of a protein. This shape-shift is the physical manifestation of information crossing a boundary. How this boundary is crossed depends entirely on the chemical nature of the ligand.
Hydrophobic vs. Hydrophilic Ligands
| Property | Behavior & Receptor Location | Examples |
|---|---|---|
| Hydrophobic Ligands | Because cell membranes are made of a lipid bilayer, hydrophobic ligands can diffuse directly across the plasma membrane. Therefore, hydrophobic ligands typically bind to intracellular receptors. | Steroid hormones are hydrophobic ligands (e.g., testosterone, estrogen). |
| Hydrophilic Ligands | Hydrophilic ligands cannot pass through the hydrophobic core of the plasma membrane. As a result, hydrophilic ligands bind to extracellular domains of cell-surface receptors. | Peptide hormones, most neurotransmitters. |
When a hydrophobic ligand crosses the membrane, it binds to a receptor inside the cell. Intracellular receptors are located in the cytoplasm or nucleus of the target cell. Once bound, these activated intracellular receptors often act as transcription factors to regulate gene expression, directly turning genes on or off.
For hydrophilic ligands stuck outside the cell, the signal must be relayed across the membrane via cell-surface receptors. There are three major classes you must know:
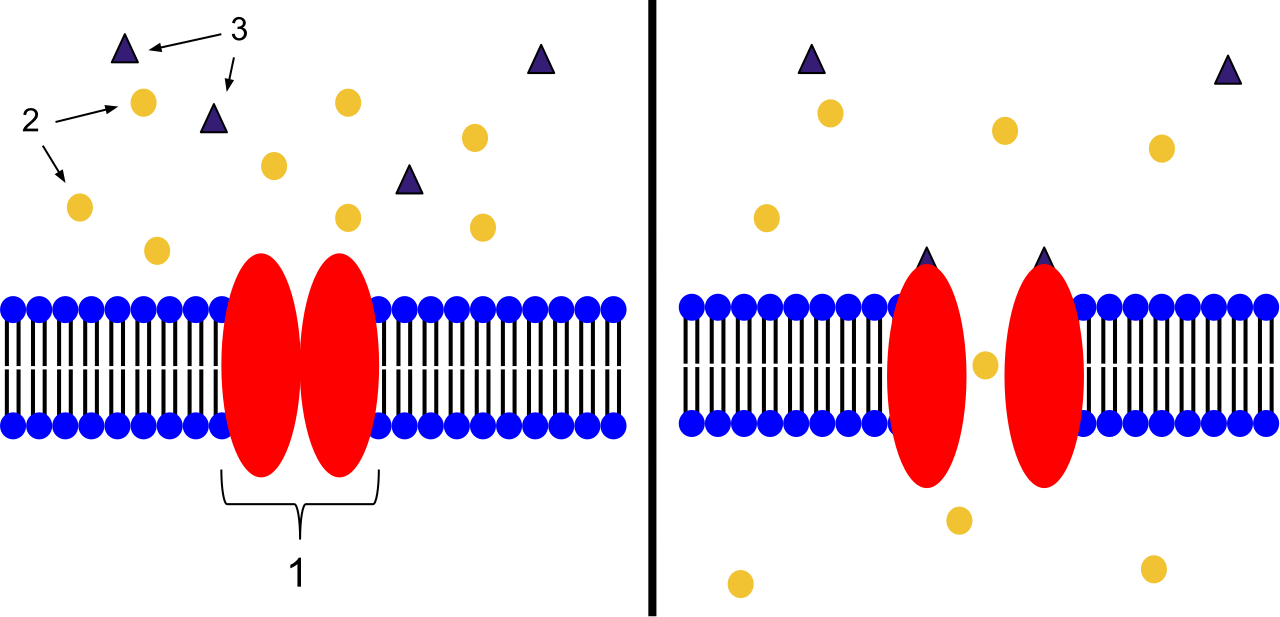
- Ion channel-linked receptors open a pore through the plasma membrane when a specific ligand binds. The conformational change physically pulls open a gateway, allowing ions (like Na+ or Ca2+) to rush in or out.
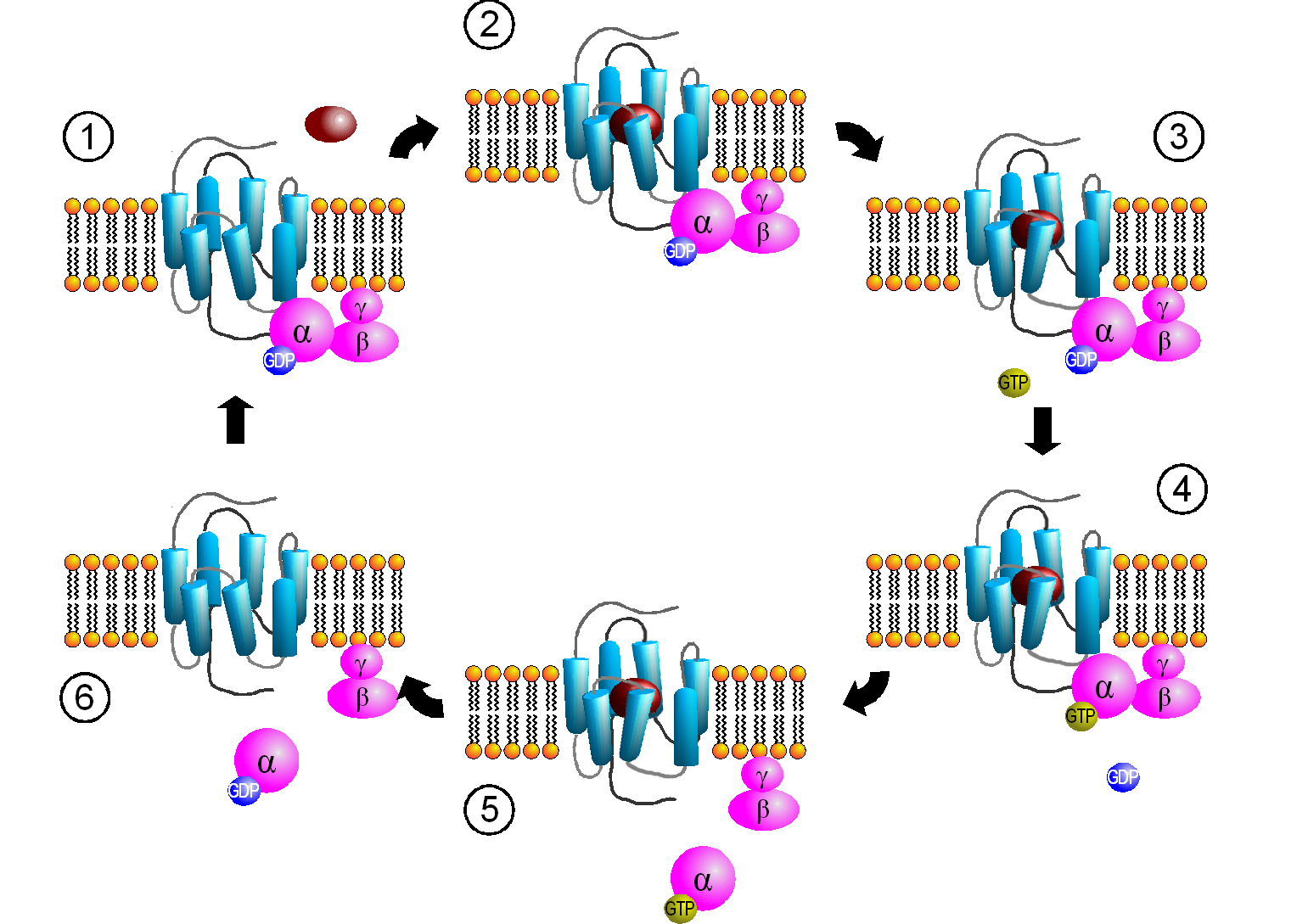
- G-protein-coupled receptors (GPCRs) bind to a ligand and activate an associated membrane protein called a G-protein. When the receptor changes shape, it physically bumps into the inactive G-protein. The result? An active G-protein exchanges bound guanosine diphosphate (GDP) for guanosine triphosphate (GTP). This swap acts as an "on" switch, allowing the G-protein to slide along the membrane and activate other enzymes.
- Enzyme-linked receptors possess intracellular domains that are associated with an enzyme. The most famous of these are Receptor Tyrosine Kinases (RTKs). Receptor tyrosine kinases typically form dimers upon ligand binding (meaning two receptor proteins pair up and lock together). Once dimerized, these receptor tyrosine kinases are enzyme-linked receptors that transfer phosphate groups to tyrosine amino acid residues on themselves and target proteins.
Stage 2: Transduction – The Whisper Down the Lane
Once the receptor has shifted shape or been phosphorylated, the message is inside the cell. But it is usually not at its final destination.
Signal transduction is the process of converting an extracellular signal into a specific intracellular response. This is rarely a single step. Instead, it is a signaling cascade—a chain of sequential biochemical events along a signal transduction pathway.
Why have a multi-step cascade instead of a direct line? Because of amplification. Signal amplification occurs when a single activated receptor activates many downstream molecules. Imagine starting a rumor. You tell three people, they each tell three people, and within minutes, the entire school knows. One single molecule of adrenaline binding to a liver cell can trigger the release of millions of glucose molecules.
The Switches: Kinases and Phosphatases
The machinery of a signaling cascade relies heavily on turning proteins "on" and "off."
- Protein kinases are enzymes that transfer phosphate groups from ATP to a target protein. Because the phosphate group is bulky and negatively charged, its addition forces the target protein to undergo a conformational change. Therefore, phosphorylation typically activates a protein within a signaling pathway.
- Conversely, protein phosphatases are enzymes that remove phosphate groups from proteins. By stripping the phosphate away, the protein returns to its resting shape. Thus, dephosphorylation typically deactivates a protein within a signaling pathway.
The Messengers
While proteins do much of the heavy lifting, cells also rely on smaller agents to rapidly broadcast signals throughout the cytoplasm. Second messengers are small, non-protein molecules or ions that relay signals inside the cell. Because they are small and water-soluble, they diffuse exceptionally quickly.
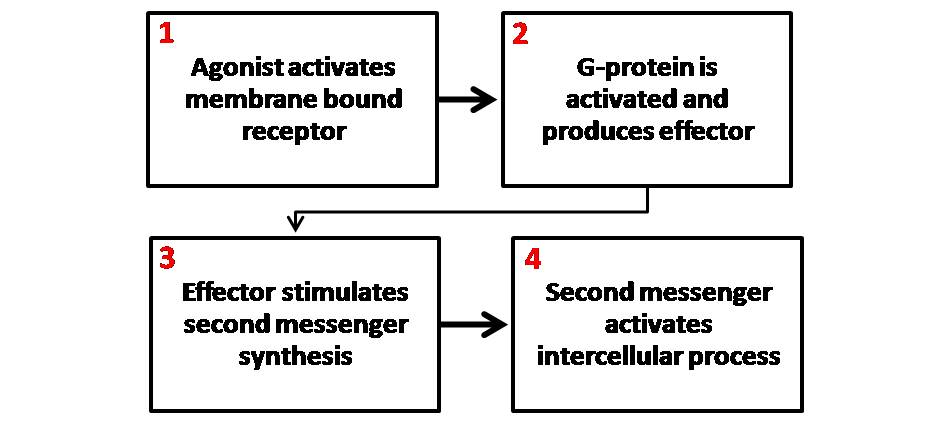
Key second messengers include:
- Cyclic AMP (cAMP): A common second messenger synthesized from ATP by the enzyme adenylyl cyclase (which is often activated by G-proteins).
- Calcium ions (Ca2+): These act as widely used second messengers in many signal transduction pathways. Cells spend a lot of energy pumping calcium out of the cytoplasm, so when a channel opens, Ca2+ floods in, acting as an intense, immediate signal.
- Inositol triphosphate (IP3) and Diacylglycerol (DAG): These are generated by the cleavage of a specific membrane lipid. IP3 is a second messenger that triggers the release of calcium ions from the endoplasmic reticulum, while DAG is a second messenger that remains in the plasma membrane to activate protein kinase C.
Stage 3: Response – The Execution of Orders
Ultimately, the cascade must result in the cell doing something. The endpoint of a signal transduction pathway is the cellular response.
Depending on the signal and the cell type, the response can vary wildly:
- A cellular response can involve changes in gene expression. (Turning on the machinery to build a new protein).
- A cellular response can involve the regulation of specific enzyme activity. (Turning on an existing enzyme to rapidly break down glycogen into glucose).
- A cellular response can trigger structural changes in the cytoskeleton. (Causing an immune cell to physically crawl toward a bacterial invader).
- In some cases, the signal tells the cell that it is time to end its lifecycle. Apoptosis is a programmed cell death triggered by specific cell signaling pathways. This is not a messy biological explosion, but a neat, highly coordinated dismantling of the cell, crucial for preventing cancer and shaping tissues during embryonic development (like removing the webbing between human fingers).
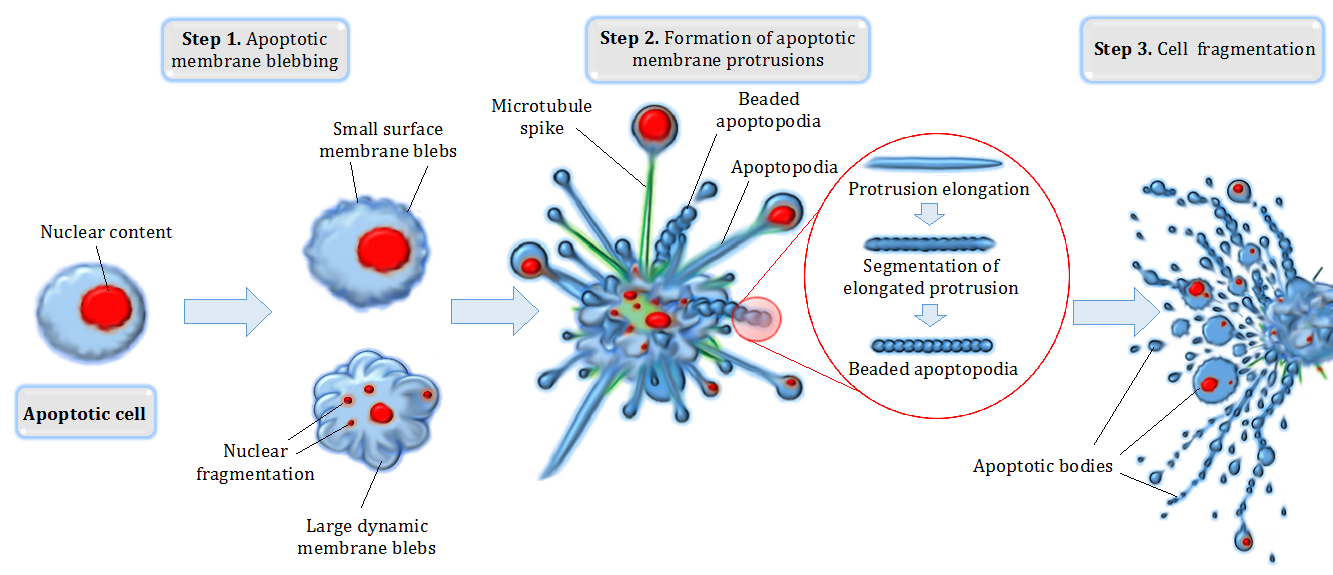
As a future educator, one of the most critical conceptual leaps you must help your students make is understanding that biological processes do not exist in a vacuum; they must be regulated. The cell must know when to stop signaling, or when to accelerate it.
Negative Feedback: The Braking System
Negative feedback mechanisms reduce the intensity of the original biological stimulus. They are self-correcting loops. If a parameter goes too high, the system pushes it back down. If it drops too low, the system pushes it back up.
Because of this stabilizing nature, negative feedback loops are the primary mechanism for maintaining homeostasis in biological systems.
Classic Example: The regulation of blood glucose levels by insulin is a classic example of a negative feedback mechanism. When you eat a heavy meal, blood glucose rises (the stimulus). The pancreas detects this and secretes insulin. Insulin acts as a ligand, binding to receptors on muscle and liver cells, causing a signaling cascade that directs the cells to absorb glucose. As blood glucose falls back to normal levels, the pancreas stops secreting insulin. The stimulus shuts off its own response.

Positive Feedback: The Accelerator
Unlike negative feedback, positive feedback mechanisms amplify the intensity of the original biological stimulus. Instead of seeking balance, they drive a process forward faster and faster. Positive feedback loops drive a specific physiological process to completion. Because they are inherently destabilizing, they are utilized only when an explosive, definitive endpoint is required.
Classic Examples:
- The release of oxytocin during childbirth is a classic example of a positive feedback mechanism. The baby's head pushes against the cervix, sending a nerve impulse to the brain. The brain releases the hormone oxytocin. Oxytocin travels through the blood and binds to receptors on the uterine muscles, causing them to contract. This pushes the baby harder against the cervix, releasing more oxytocin, causing stronger contractions, until the baby is born.
- Blood clotting cascades are regulated by positive feedback mechanisms. Once a blood vessel is damaged, activated platelets release chemical signals that attract and activate more platelets, rapidly building a clot to seal the wound before the organism bleeds out.
- In agriculture and botany, ethylene gas production during fruit ripening is an example of a positive feedback mechanism in plants. As one apple begins to ripen, it releases ethylene gas (a paracrine and endocrine-like signal). This gas triggers neighboring apples to ripen, causing them to release their own ethylene. This is why "one bad apple spoils the bunch."
When you design your future lesson plans on cell communication, ground the invisible chemistry in the highly visible logic of cause and effect. A ligand has a shape. A receptor responds to that shape. The resulting cascade amplifies the signal, utilizing kinases and second messengers, resulting in a physical response—all of which is governed by the beautiful, mathematical logic of feedback loops. Master this narrative, and you will not only ace your Content Knowledge exam; you will demystify the microscopic world for thousands of students over your career.