Cell Theory and Organization
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
If you take a thin slice of plant tissue and place it beneath a simple lens, the illusion of solid matter shatters. You find instead a microscopic honeycomb of walled chambers. Robert Hooke first made this observation in 1665 after observing cork tissue under a microscope, famously coining the term "cell" to describe these structures. Though he was looking at the skeletal remains of dead plant matter, Hooke’s observation initiated the greatest unifying principle in biology: the realization that the bewildering diversity of life is built from the exact same fundamental architectural unit.
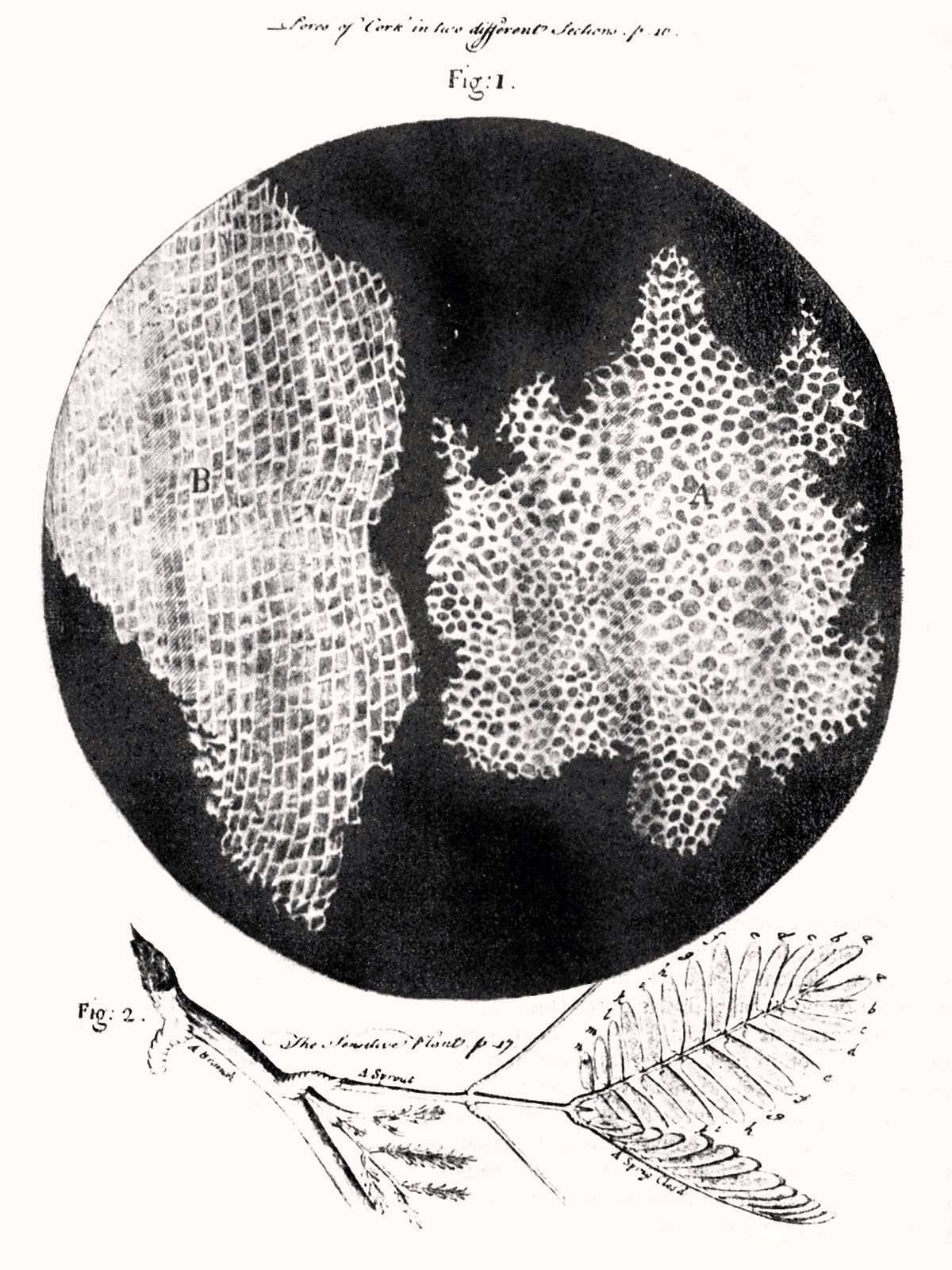
For you as a future biology educator, the cell is where the story of life truly begins. It is the boundary line between the chaotic chemistry of the nonliving universe and the highly ordered machinery of life.
When your students ask, "What makes something alive?", they are asking a question that has consumed biologists for centuries. We define life not by a single essence, but by a collection of functional characteristics. To be considered alive, an entity must possess all of them.
Living things exhibit highly structured cellular organization. They require continuous energy input to perform metabolic processes. They do not merely inflate; living things grow and develop following specific instructions coded by genetic material. They reproduce to pass genetic information to the next generation. They respond dynamically to environmental stimuli, and they maintain internal stability through physiological homeostasis. Over generations, populations of living things evolve over time through natural selection and adaptation.
This strict biological rubric provides a clear boundary. Viruses are classified as nonliving entities by most biological standards. Why? Because viruses cannot perform metabolic functions or reproduce outside of a host cell. A virus on a doorknob is as inert as a grain of salt until it hijacks the machinery of a living cell.
The Evolution of Cell Theory
The realization that all life is cellular did not happen overnight. After Hooke, Anton van Leeuwenhoek was the first scientist to observe living cells under a microscope, marveling at the microscopic "animalcules" swimming in pond water. Centuries later, Matthias Schleiden proposed that all plant tissues are composed of cells, and Theodor Schwann proposed that all animal tissues are composed of cells. Finally, Rudolf Virchow popularized the concept that all cells come from pre-existing cells.
Together, these observations formed the classical Cell Theory:
- All living organisms are composed of one or more cells.
- The cell is the basic unit of structure and organization in all living organisms.
- All cells arise from pre-existing cells through cellular division.
As biochemical techniques improved, the classical theory was expanded. Modern cell theory states that:
- Energy flow occurs within cells.
- Genetic information is passed on from cell to cell during division.
- All cells share the same basic chemical composition.
A common question in the biology classroom is, "If cells are so efficient, why aren't humans just one giant cell?"
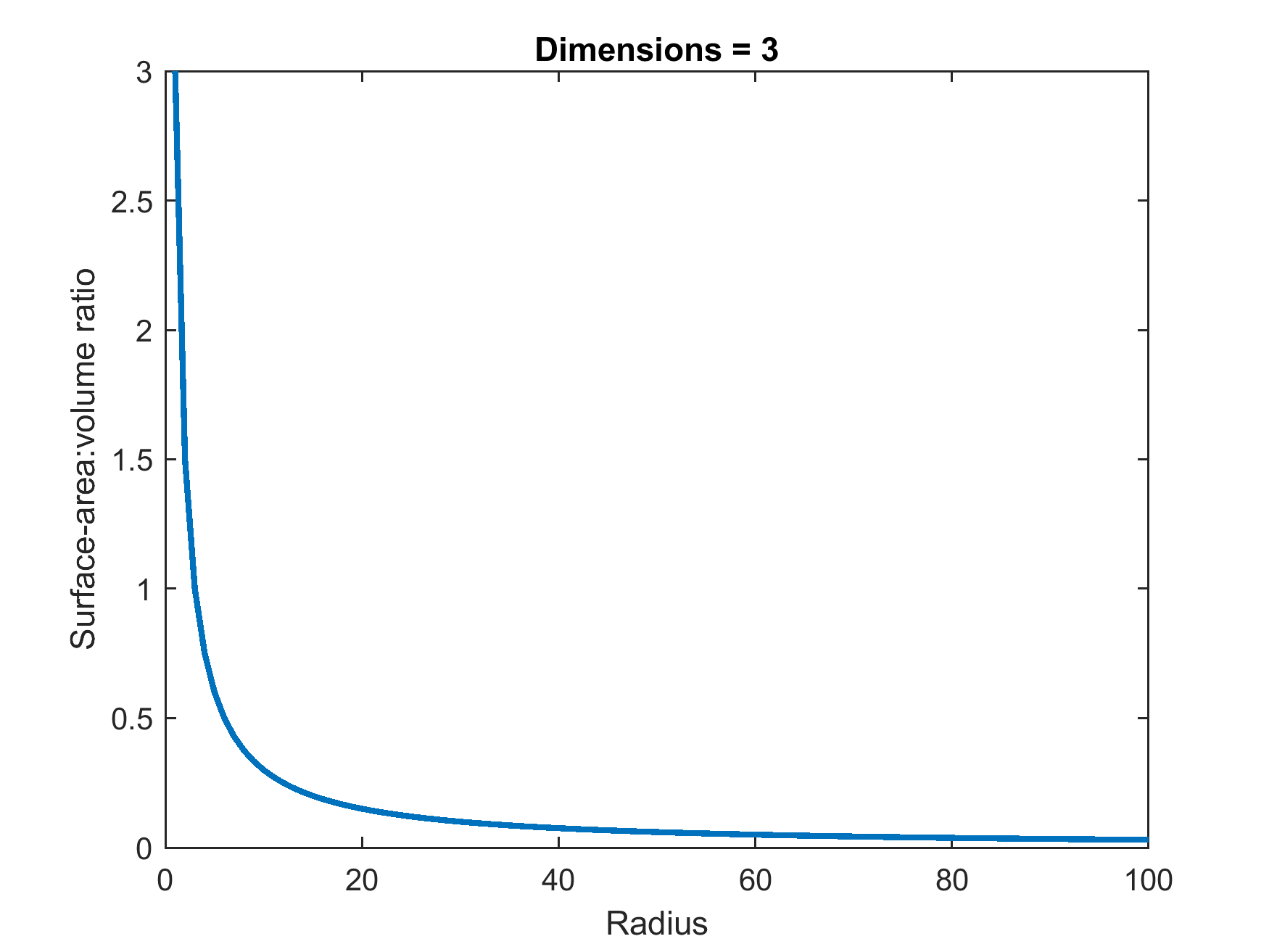
The answer lies in geometry. The surface area-to-volume ratio physically limits the maximum functional size a cell can achieve. As a cell increases in size, the cellular volume increases at a faster rate than the cellular surface area.
Think of the cell membrane as a border crossing. All nutrients entering the cell, and all waste leaving it, must pass through this membrane. If the internal volume (the bustling city) grows too massive, the surface area (the border) cannot accommodate the necessary traffic. A decreasing surface area-to-volume ratio severely reduces the efficiency of nutrient uptake across the cell membrane. Therefore, life builds complex organisms by multiplying cells, not by perpetually expanding a single one.
Despite the immense variety of life, all living cells possess a plasma membrane, cytoplasm, DNA, and ribosomes. From there, cellular architecture diverges into two fundamental plans: prokaryotic and eukaryotic.
The Prokaryotic Cell
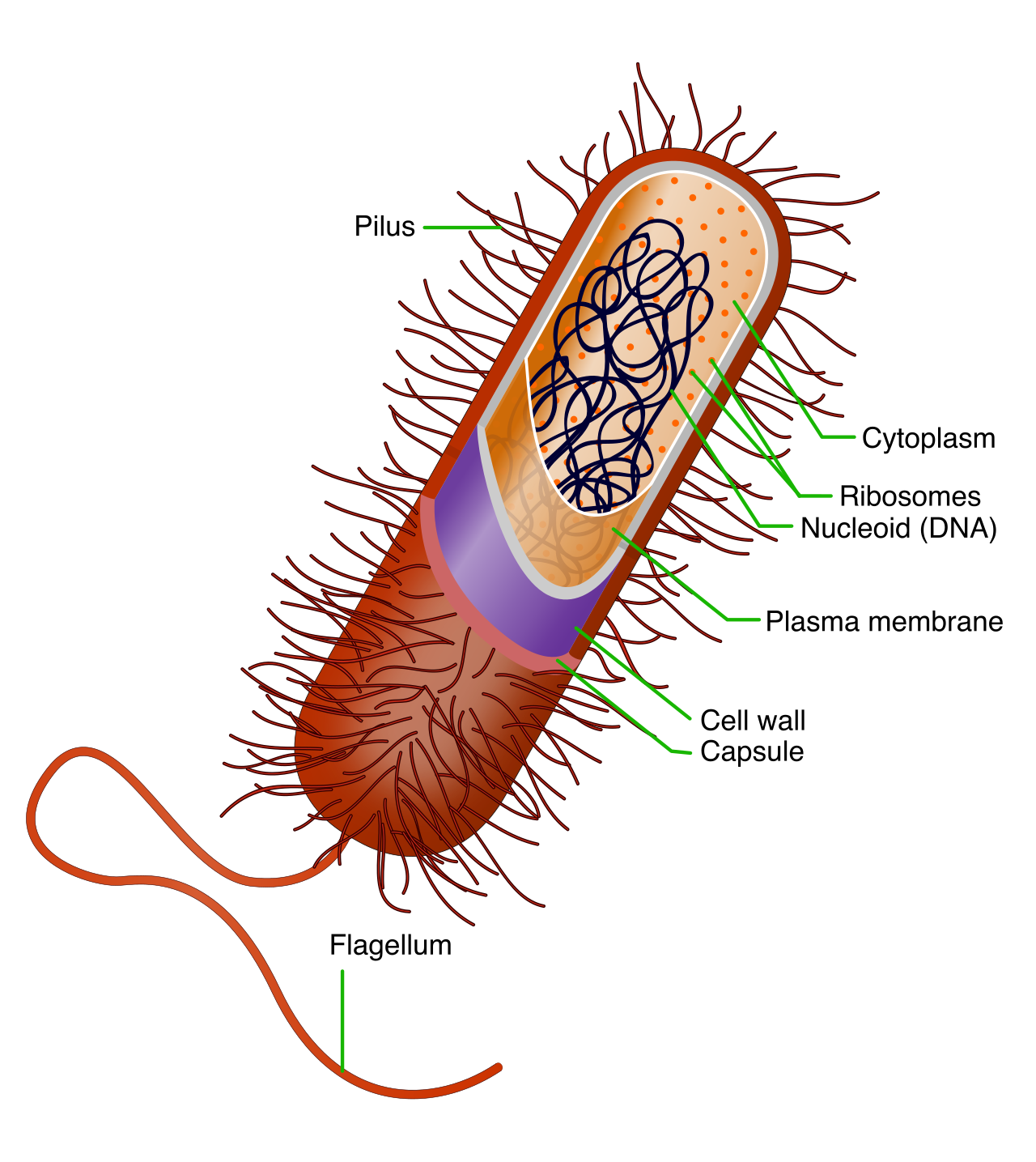
Organisms in the domains Bacteria and Archaea are exclusively classified as prokaryotes. Prokaryotic cells are built for rapid, streamlined survival. They are generally significantly smaller than eukaryotic cells, typically ranging from 0.1 to 5.0 micrometers in diameter.
Prokaryotic cells completely lack a membrane-bound nucleus and strictly lack membrane-bound organelles. Instead, prokaryotic genetic material is localized within an irregularly shaped region called the nucleoid. Most prokaryotic genomes consist of a single, circular double-stranded DNA molecule. In addition to this main chromosome, many bacteria carry plasmids—small, circular accessory DNA molecules naturally found in many prokaryotic organisms. Crucially, plasmids replicate entirely independently of the main chromosomal DNA in prokaryotes, often carrying beneficial genes like those for antibiotic resistance.
Because they are so compact, the small size of prokaryotic cells allows ions and organic molecules to rapidly diffuse to all parts of the cell. Many bacterial cells possess a rigid external cell wall primarily composed of peptidoglycan, giving them shape and protection. To move through fluid environments, prokaryotic flagella act as rigid rotary motors to propel the cell forward. Finally, the protein factories of the prokaryote, its ribosomes, have a sedimentation coefficient of 70S.
The Eukaryotic Cell
Animals, plants, fungi, and protists are exclusively classified as eukaryotes. Eukaryotic cells are much larger, typically ranging from 10 to 100 micrometers in diameter.
Eukaryotic cells contain a distinct membrane-bound nucleus that houses the primary genetic material. Unlike bacteria, eukaryotic nuclear chromosomes are typically linear double-stranded DNA molecules. Deep within this nucleus sits the nucleolus, a dense region within the eukaryotic nucleus dedicated to synthesizing ribosomal RNA. Eukaryotic cytoplasmic ribosomes are larger than their prokaryotic counterparts, with a sedimentation coefficient of 80S.
The hallmark of the eukaryote is its specialized membrane-bound organelles such as mitochondria and the endoplasmic reticulum. This organelle compartmentalization in eukaryotic cells allows incompatible chemical reactions to occur simultaneously in isolated environments. However, the larger volume of eukaryotic cells necessitates specialized intracellular transport systems to move molecules efficiently—diffusion alone is no longer fast enough. When eukaryotes need to swim, their eukaryotic flagella utilize a flexible, whip-like undulating motion to propel the cell through fluid environments.
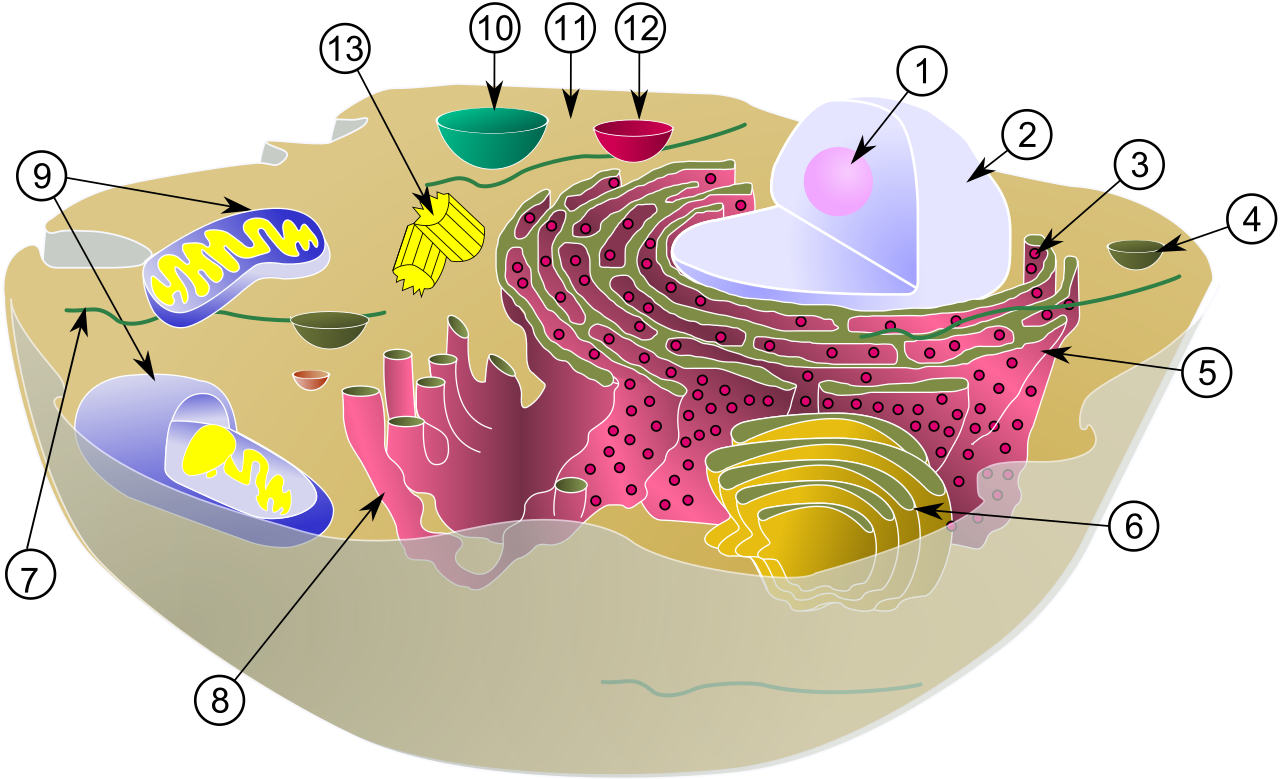
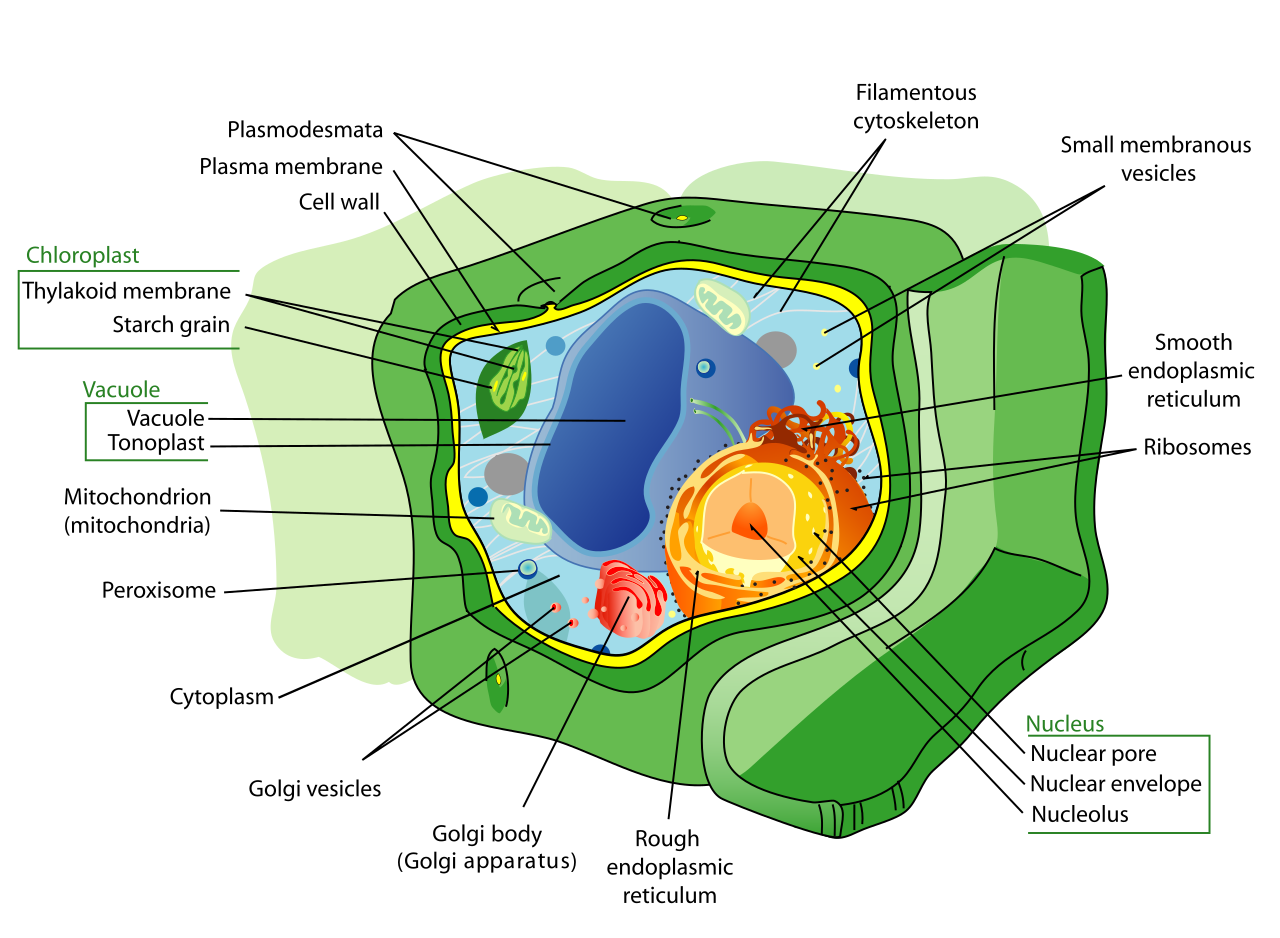
Plant vs. Animal Cells
Within the eukaryotic domain, plant and animal cells have distinct functional differences that you will teach extensively.
- Boundary: Plant cells possess a rigid external cell wall primarily composed of cellulose. Animal cells lack a rigid external cell wall, allowing for the flexible variety of cell types (like muscle and nerve cells) found in animals.
- Energy: Plant cells contain chloroplasts responsible for carrying out photosynthesis. Animal cells naturally lack chloroplasts.
- Storage: Plant cells typically feature a large central vacuole that maintains internal turgor pressure. When a plant is well-watered, this vacuole pushes against the rigid cell wall, keeping the plant upright.
| Feature | Prokaryotes | Eukaryotes |
|---|---|---|
| Size | 0.1 to 5.0 µm | 10 to 100 µm |
| Nucleus | Absent (Nucleoid region) | Present (Membrane-bound) |
| DNA | Single, circular double-stranded | Linear double-stranded |
| Organelles | Absent | Present (Mitochondria, ER, etc.) |
| Ribosomes | 70S | 80S |
| Flagella | Rigid rotary motor | Flexible, whip-like |
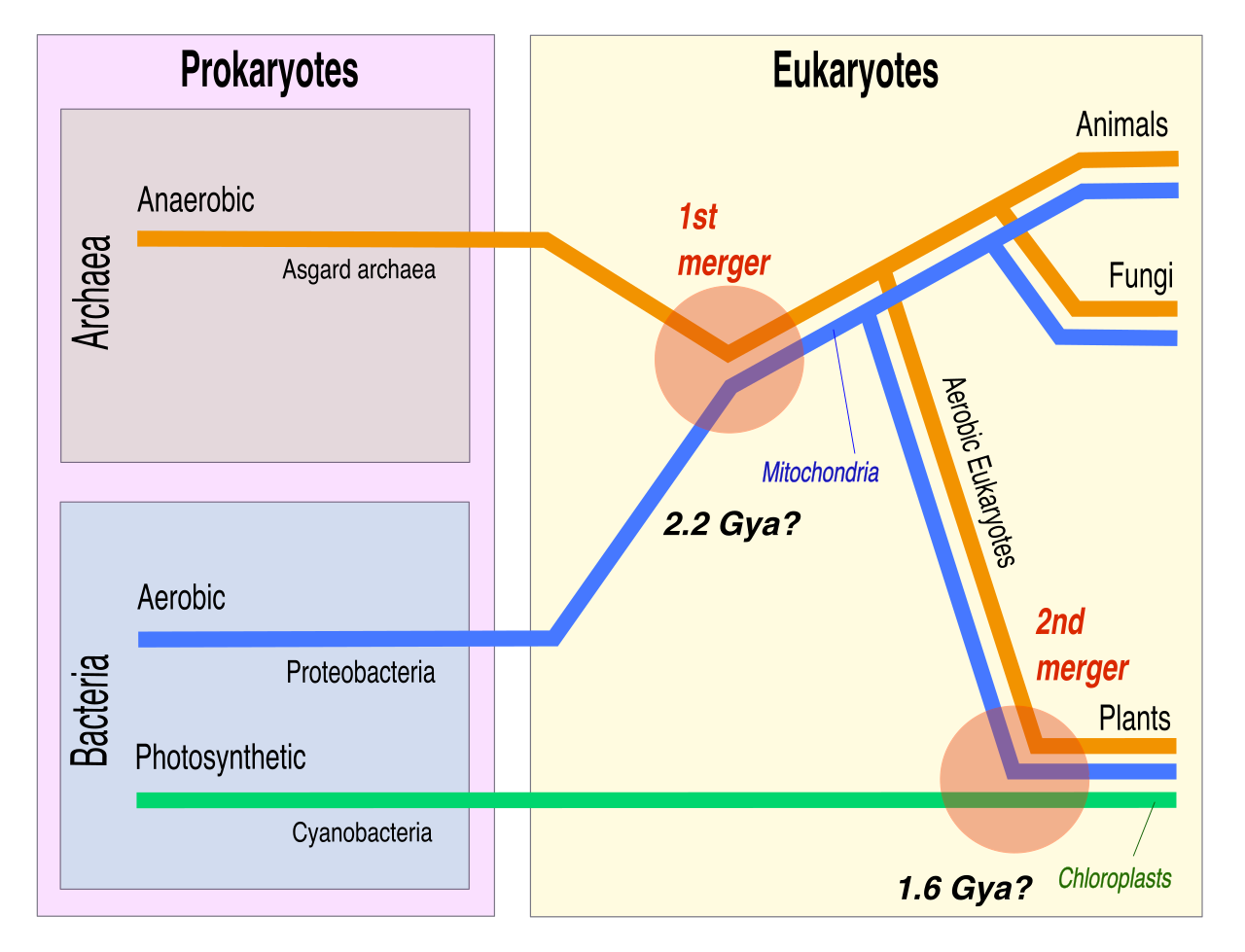
One of the most fascinating narratives in evolutionary biology is the origin of eukaryotic organelles. How did the complex eukaryotic cell acquire its energy-producing structures?
The endosymbiotic theory proposes that mitochondria originated from engulfed free-living aerobic prokaryotes. Millions of years ago, an ancestral cell consumed an oxygen-using bacterium, but instead of digesting it, formed a symbiotic relationship. Similarly, the endosymbiotic theory proposes that chloroplasts originated from engulfed free-living photosynthetic prokaryotes.
The evidence for this is elegantly simple and structurally undeniable: both mitochondria and chloroplasts contain independent circular DNA distinct from the host cell nuclear genome. They even possess their own 70S ribosomes and replicate much like modern bacteria do, whispering to their ancient, independent past.
A cell is not a static water balloon; it is a highly active machine constantly adjusting to its surroundings. Homeostasis is the active maintenance of a stable internal environment despite external fluctuations.
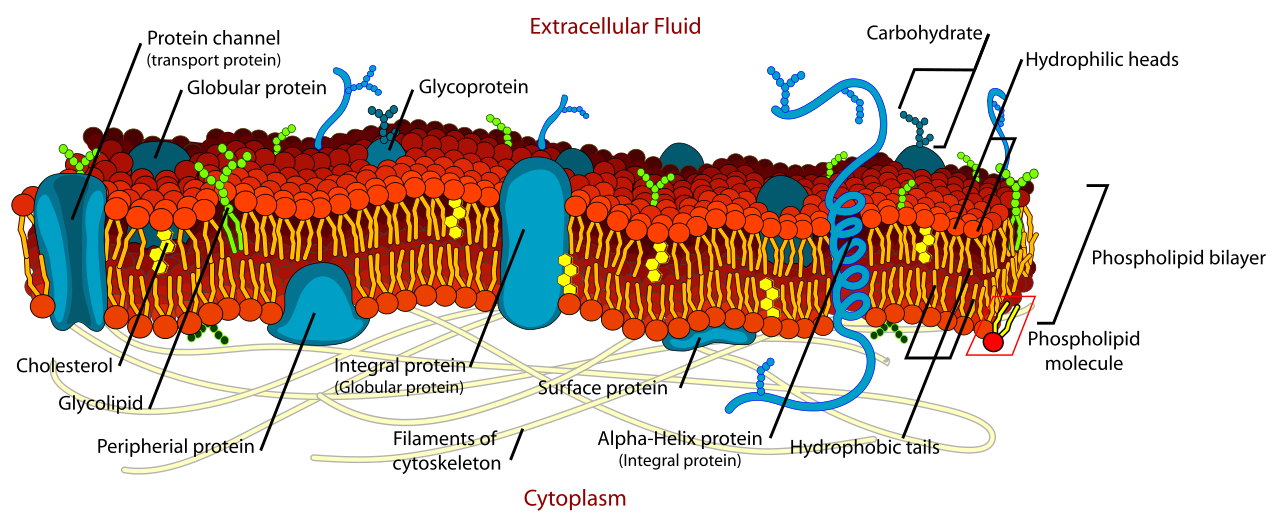
To maintain this stability, the plasma membrane regulates the passage of specific substances to maintain cellular homeostasis. This selective permeability allows cells to precisely control internal concentrations of ions and molecules. It achieves this via distinct transport mechanisms:
- Passive Transport: Cells use passive transport to move substances down a concentration gradient without energy input. It is the biological equivalent of rolling a boulder down a hill.
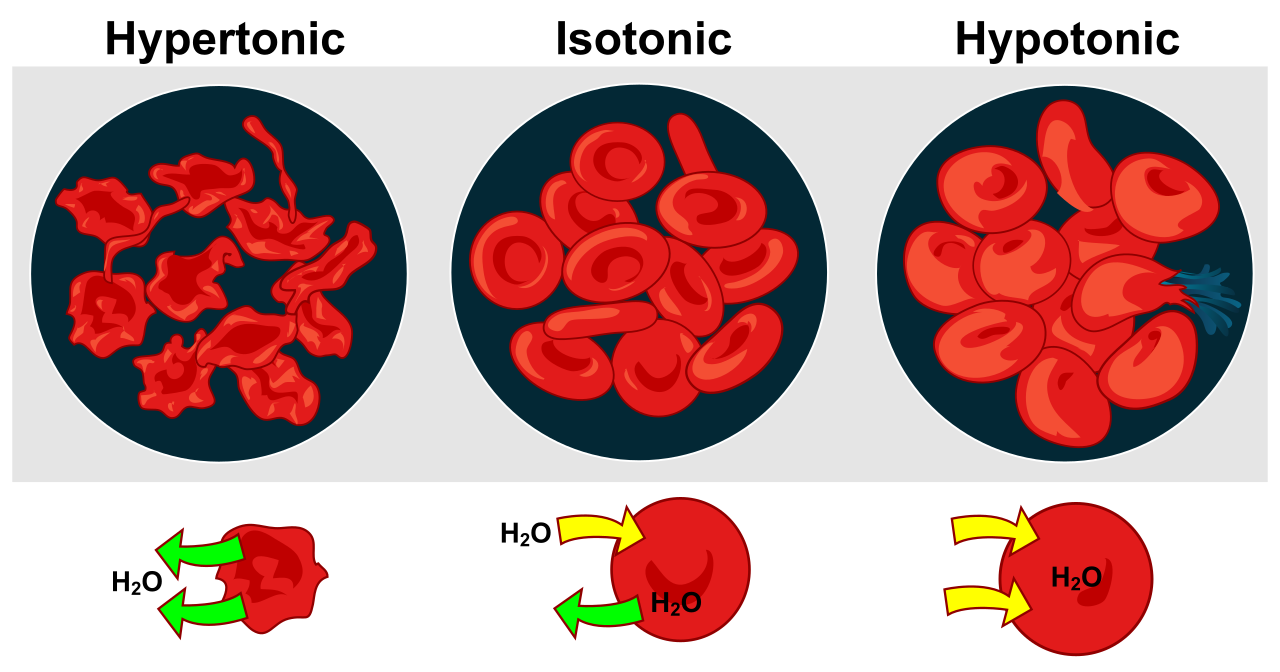
- Osmosis: A specific type of passive transport, osmosis is the spontaneous diffusion of water across a semipermeable membrane to balance solute concentrations. Water will move to where solutes are more highly concentrated.
- If a cell is placed in a hypotonic environment (where the surrounding fluid has fewer solutes than the cell interior), cells respond to hypotonic environments by taking in water.
- Conversely, cells respond to hypertonic environments by losing water to the surroundings, often leading to cellular shrinkage.
- Active Transport: Sometimes a cell needs to stockpile a nutrient or expel a waste product against natural diffusion. Cells use active transport mechanisms to move substances against a concentration gradient. Because this is like pushing a boulder up a hill, active transport requires the direct expenditure of cellular energy in the form of ATP.
Cellular Communication and Feedback
Cells do not exist in isolation; they continuously monitor their external environment. Cells rely on specific membrane receptor proteins to detect chemical signals in the external environment. When a molecule binds to a receptor on the outside of the cell, it sets off a domino effect inside. These signal transduction pathways convert extracellular signals into distinct intracellular responses, telling the cell to divide, secrete a hormone, or undergo programmed death.
Finally, at the whole-organism level, living things rely on internal regulation to survive. Negative feedback loops counter deviations from a target set point to restore cellular equilibrium. If a cell becomes too hot, it initiates processes to cool down; once the normal temperature is reached, the cooling process is shut off.

Understanding these mechanisms is vital. When your students understand how a cell uses its membrane, transports its resources, and communicates via feedback loops, they cease to see biology as a list of vocabulary words. They begin to see it as the grand, dynamic engineering of life itself.