Photosynthesis
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Hold a heavy block of solid oak wood in your hands. It feels dense, permanent, and massive. A profound conceptual hurdle for any biology student—and one of the most magnificent realities you will convey as a teacher—is the realization that almost all of that physical mass came from invisible gas. Trees are built from thin air. The machinery that accomplishes this feat is arguably the most important biological process on Earth, bridging the physics of electromagnetism with the chemistry of carbon.
As a biology teacher, your goal is to demystify this process so your students see the leaf not just as a green biological ornament, but as a hyper-efficient, solar-powered chemical factory. We must break down how light is harvested, how water is shattered, and how a gas is knit together into the sugars that form the foundation of global food webs.
Here is the definitive guide to the machinery of photosynthesis.
At its core, photosynthesis is the process by which photoautotrophs convert light energy into chemical energy. To teach this effectively, you must first orient your students to the overall reaction and the physical space where it occurs.
The Grand Equation: 6CO2+6H2O+light energy→C6H12O6+6O2
In this equation, we see the inputs (carbon dioxide, water, and photons) and the outputs (glucose and oxygen). But how do the inputs get in? Plant leaves exchange carbon dioxide and oxygen through microscopic epidermal pores called stomata.
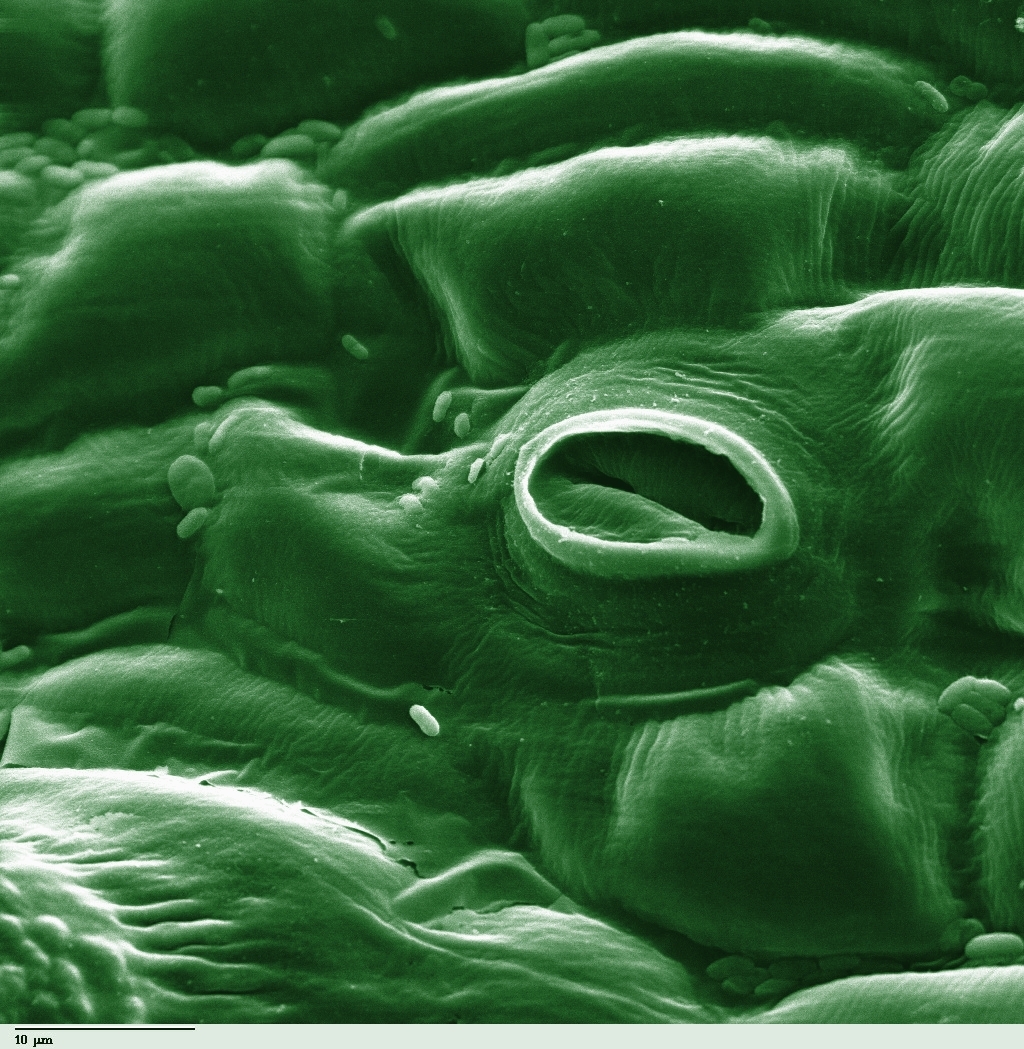
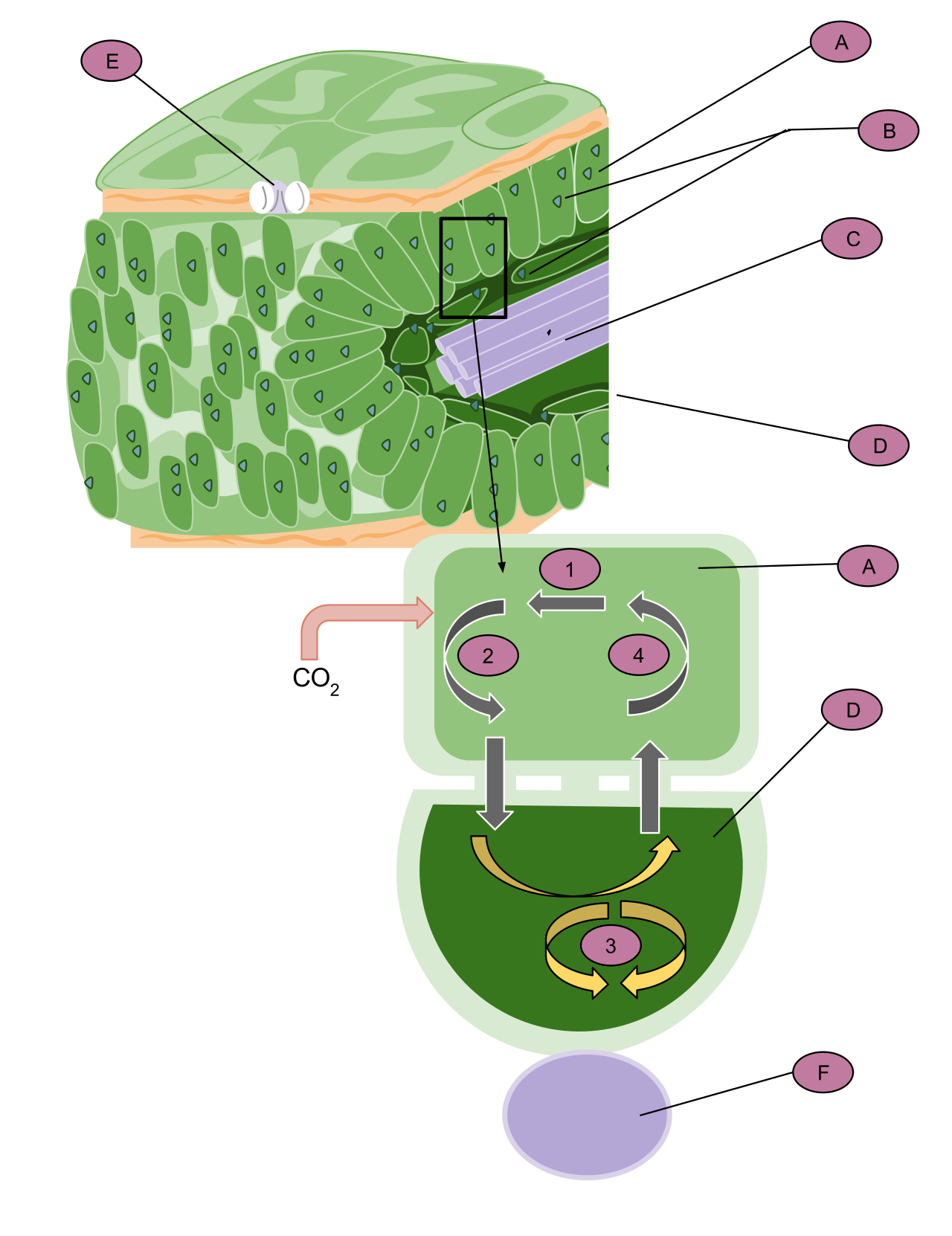
Once inside the cell, photosynthesis in eukaryotes takes place within specialized organelles called chloroplasts.
If you slice open a chloroplast, you don't find an empty sac. You find a highly organized internal structure. Chloroplasts contain a third, inner membrane system called the thylakoid membrane. These thylakoids are stacked into disc-like structures called grana (singular: granum), looking much like stacks of microscopic green coins.
The architecture establishes two distinct zones where the two major phases of photosynthesis occur:
- The Thylakoid Lumen: The continuous internal space enclosed by the thylakoid membrane.
- The Stroma: The aqueous fluid surrounding the thylakoids inside the chloroplast.
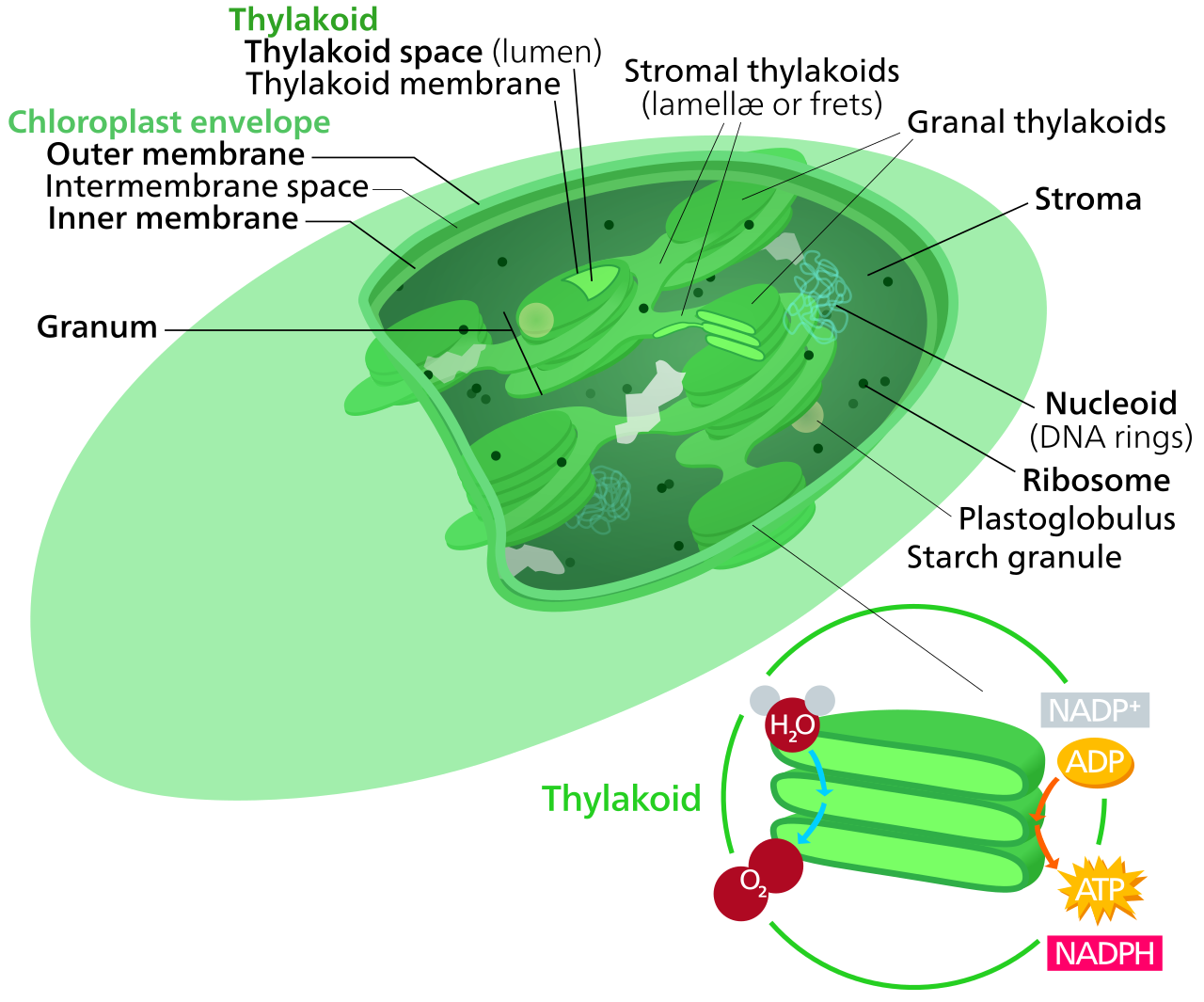
The geography dictates the chemistry: the light-dependent reactions of photosynthesis occur within and across the thylakoid membrane, while the sugar-building Calvin-Benson cycle occurs in the stroma of the chloroplast.
The purpose of this first phase is to trap the kinetic energy of sunlight and package it into transient chemical batteries. Specifically, the light-dependent reactions capture light energy to synthesize ATP and NADPH.
Capturing the Photon
Embedded within the thylakoid membrane are remarkable molecular machines known as photosystems. Photosystems consist of a light-harvesting complex and a reaction center.
Think of a photosystem as a giant satellite dish. Chlorophyll molecules absorb photons of light to excite electrons to a higher energy state. The light-harvesting complex funnels resonance energy from absorbed light to the reaction center, focusing the energy into a highly specific pair of chlorophyll molecules.
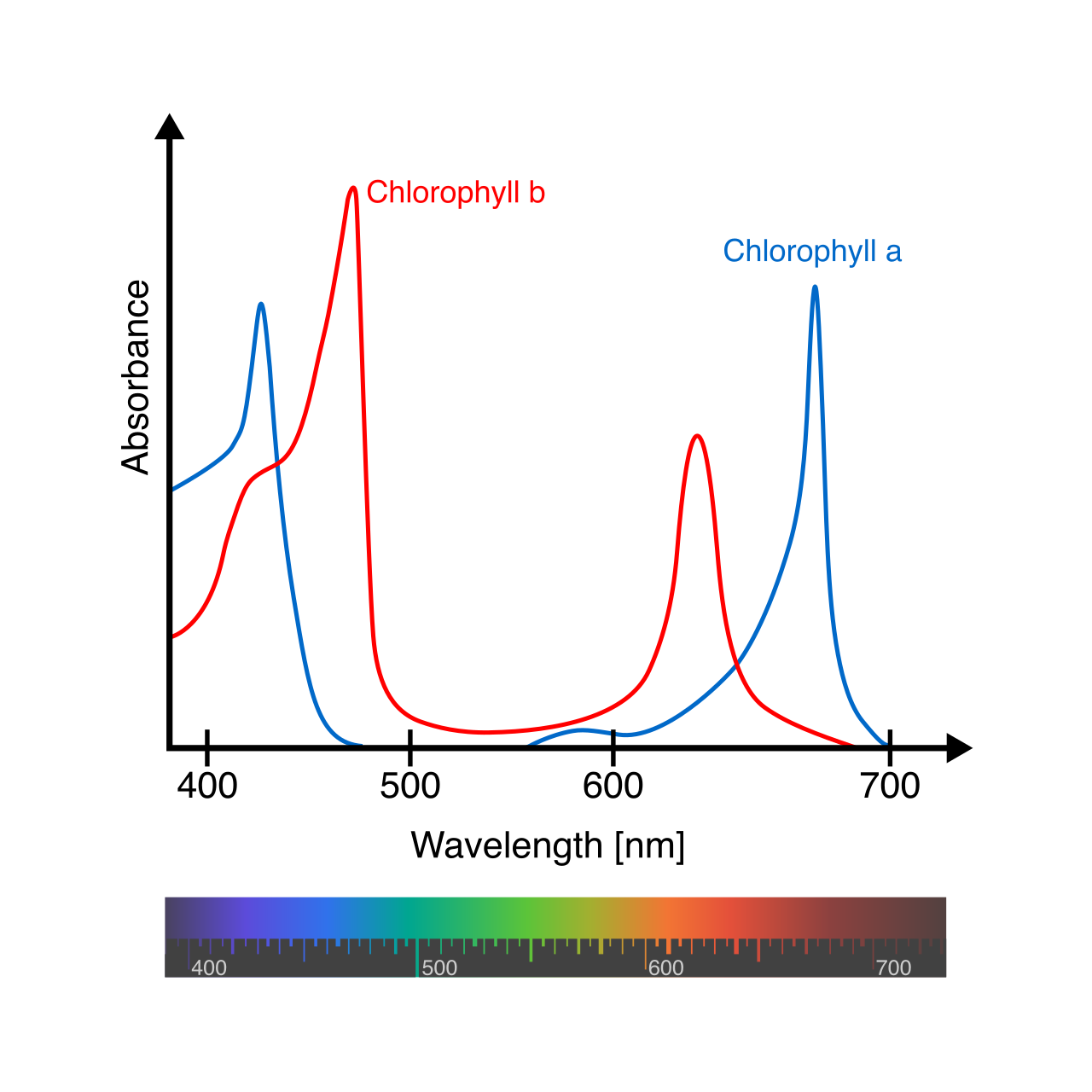
Photosystem II and the Splitting of Water
The journey of an electron begins in Photosystem II, which is a protein complex in the thylakoid membrane containing a special pair of chlorophyll a molecules called P680 (named because they best absorb light at a wavelength of 680 nanometers).
When the funneled energy reaches P680, an electron becomes so energized that it physically leaves the molecule: an excited electron from the P680 reaction center in Photosystem II is transferred to a primary electron acceptor.
But now, Photosystem II has a problem. It is missing an electron. It is positively charged and highly electronegative. To survive, Photosystem II replaces its lost electrons by catalyzing the splitting of water molecules.
Photolysis: The light-driven splitting of water molecules. Photolysis in Photosystem II produces oxygen gas, protons (H+), and electrons.
This is where the oxygen we breathe originates! It is simply the exhaust gas of a molecular machine desperate to replace its stripped electrons.
The Electron Transport Chain and Chemiosmosis
The excited electron doesn't just wander off. Electrons travel from Photosystem II to Photosystem I through a thylakoid electron transport chain (ETC).
As the electron falls to a lower energy state moving through the ETC, that released energy is put to work. The electron transport chain actively pumps protons from the stroma into the thylakoid lumen. Because the thylakoid lumen is a small, enclosed space, the accumulation of protons in the thylakoid lumen creates an electrochemical proton gradient.
Here is where you use the classic "hydroelectric dam" analogy for your students. Water trapped behind a dam holds immense potential energy. When allowed to flow through a turbine, it generates electricity. Similarly, the process of generating ATP using a proton gradient across a membrane is called chemiosmosis.
The "turbine" in the thylakoid membrane is an enzyme. ATP synthase uses the electrochemical proton gradient to phosphorylate ADP into ATP as protons rush back out into the stroma.
Photosystem I and NADPH Synthesis
By the time the electron reaches the end of the chain, it is exhausted. Enter the second satellite dish: Photosystem I, a protein complex in the thylakoid membrane containing a special pair of chlorophyll a molecules called P700.
Photosystem I re-energizes depleted electrons from the electron transport chain by absorbing additional light energy. Now highly energetic once more, these energized electrons from Photosystem I are transferred to the enzyme NADP+ reductase.
Finally, NADP+ reductase catalyzes the transfer of electrons to NADP+ to form the electron carrier NADPH.
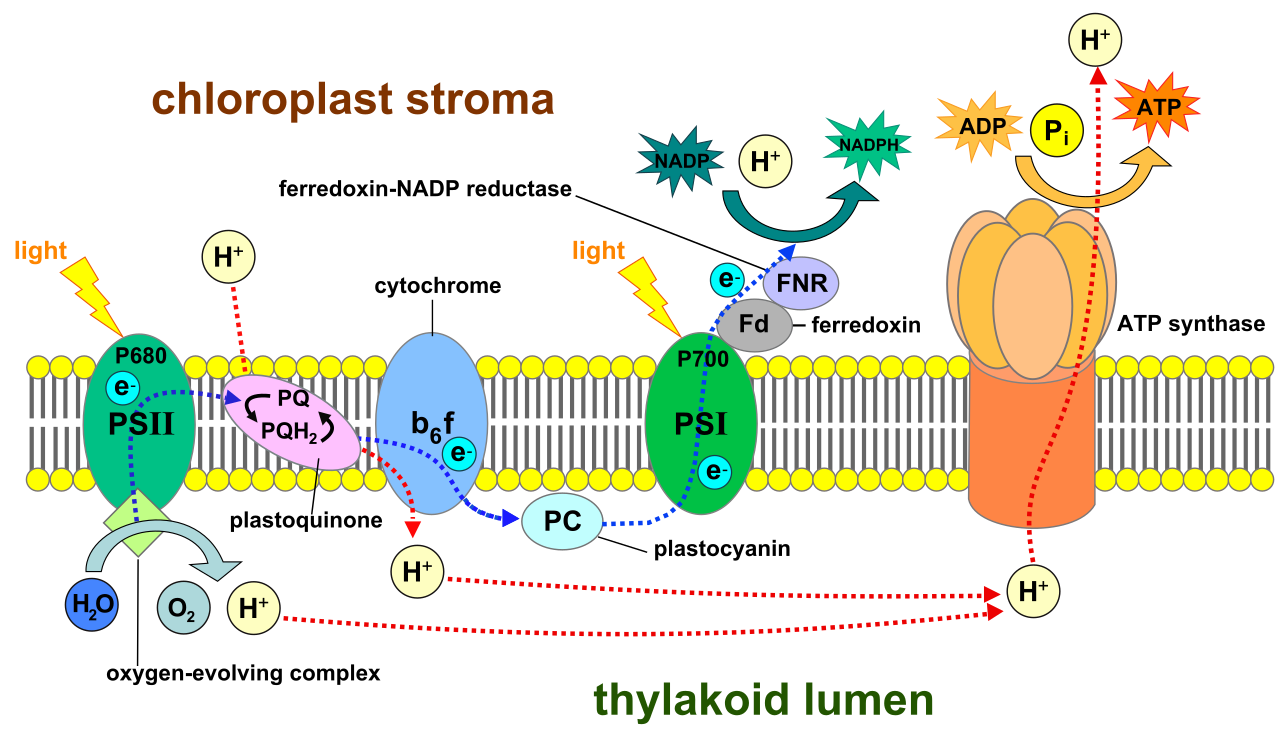
Linear vs. Cyclic Flow
What we just walked through is the standard pathway: Linear electron flow utilizes both photosystems and produces ATP, NADPH, and oxygen.
However, chloroplasts are dynamic. Sometimes, the cell needs more ATP but has plenty of NADPH. In this case, it shifts gears. Cyclic electron flow involves only Photosystem I and produces ATP without producing NADPH or oxygen. Electrons simply cycle out of PSI, fall down the proton-pumping ETC to generate a gradient, and return right back to PSI.
Now we have our fully charged batteries—ATP and NADPH—waiting in the stroma. It is time to build the wood from the air.
The Calvin-Benson cycle uses ATP and NADPH to reduce carbon dioxide into sugar molecules. This cycle, occurring in the stroma, operates as a relentless molecular assembly line. The Calvin-Benson cycle consists of three major phases: carbon fixation, reduction, and regeneration.
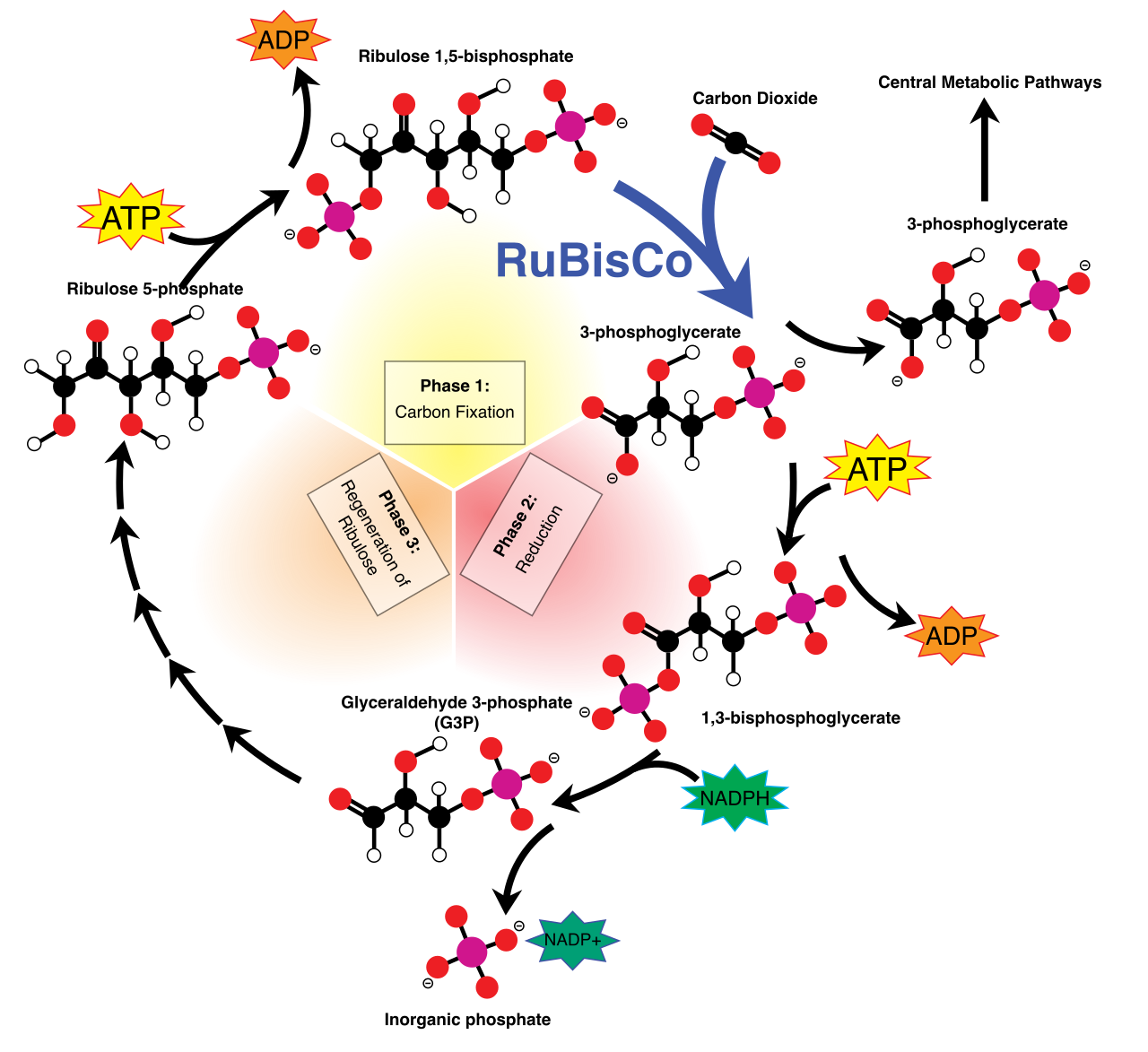
Phase 1: Carbon Fixation
The cycle begins by capturing inorganic gas. Ribulose-1,5-bisphosphate (RuBP) is a five-carbon sugar molecule utilized as the carbon dioxide acceptor in the Calvin-Benson cycle.
The critical enzyme here is RuBisCO, arguably the most abundant protein on Earth. RuBisCO is the enzyme that catalyzes the attachment of carbon dioxide to ribulose-1,5-bisphosphate.
When you fuse a 1-carbon CO2 to a 5-carbon RuBP, you get 6 carbons. The reaction between carbon dioxide and ribulose-1,5-bisphosphate produces a highly unstable six-carbon intermediate. Because it is so unstable, it violently and immediately splits into two molecules of 3-phosphoglycerate (3-PGA), which are 3-carbon molecules.
Phase 2: Reduction
The 3-PGA molecules are effectively organic, but they are stuck at a low energy state. We need to "reduce" them (add high-energy electrons). During the reduction phase, ATP and NADPH are used to convert 3-phosphoglycerate into glyceraldehyde 3-phosphate (G3P).
Glyceraldehyde 3-phosphate (G3P) is a three-carbon sugar phosphate produced by the Calvin-Benson cycle. It is the ultimate payload of photosynthesis—the raw material from which plants synthesize glucose, cellulose (wood), and starch.
Phase 3: Regeneration
If the plant used all the G3P to make glucose, the cycle would grind to a halt because it would run out of RuBP to accept the next CO2. Therefore, one molecule of glyceraldehyde 3-phosphate exits the Calvin-Benson cycle for every three molecules of carbon dioxide fixed.
The remaining molecules stay behind to keep the wheel turning. Through a complex series of rearrangements, five molecules of glyceraldehyde 3-phosphate are utilized to regenerate three molecules of ribulose-1,5-bisphosphate. This spatial reorganization takes energy; the regeneration of ribulose-1,5-bisphosphate requires the hydrolysis of additional ATP molecules.
The Energetic Cost (The Stoichiometry)
To help your future students grasp this, frame it as an energetic budget:
- The synthesis of one net molecule of glyceraldehyde 3-phosphate requires three turns of the Calvin-Benson cycle.
- Because glucose is a 6-carbon molecule and G3P is a 3-carbon molecule, the synthesis of one glucose molecule requires six complete turns of the Calvin-Benson cycle.
- To fuel this math, six turns of the Calvin-Benson cycle consume 18 ATP molecules and 12 NADPH molecules.
Biology is wondrous, but it is not perfectly engineered. RuBisCO, the crucial enzyme of carbon fixation, has a glaring structural flaw. It evolved billions of years ago when the atmosphere had almost no oxygen. Therefore, photorespiration occurs when the enzyme RuBisCO binds oxygen instead of carbon dioxide.
When this happens, the plant enters a metabolic dead-end. Photorespiration consumes ATP and releases carbon dioxide without producing functional sugar molecules. It is a massive drain on the plant's resources. On hot, dry days, stomata close to prevent water loss, oxygen builds up inside the leaf, and photorespiration skyrockets.
To survive this, evolution devised two brilliant workarounds:
- C4 plants minimize photorespiration by physically separating initial carbon fixation in mesophyll cells from the Calvin cycle in bundle-sheath cells. (A spatial solution: they pump carbon into a deep interior vault where RuBisCO is kept away from oxygen).
- CAM plants minimize photorespiration by temporarily separating initial carbon fixation at night from the Calvin cycle during the day. (A temporal solution: cacti and succulents open their stomata only at night to bank carbon, running the Calvin cycle behind closed doors during the hot day).
While you will spend most of your time teaching photosynthesis, your students must understand that sunlight is not the only way to power an ecosystem.
Chemosynthesis is the biological process of synthesizing organic compounds using energy derived from the oxidation of inorganic chemicals.
Because there is no sun in the abyssal zone of the ocean, many chemosynthetic organisms are found in extreme environments like deep-sea hydrothermal vents. Unlike plants, chemosynthetic organisms rely on inorganic electron donors such as hydrogen sulfide, ammonia, or methane, meaning chemosynthesis does not require light energy to produce organic molecules.
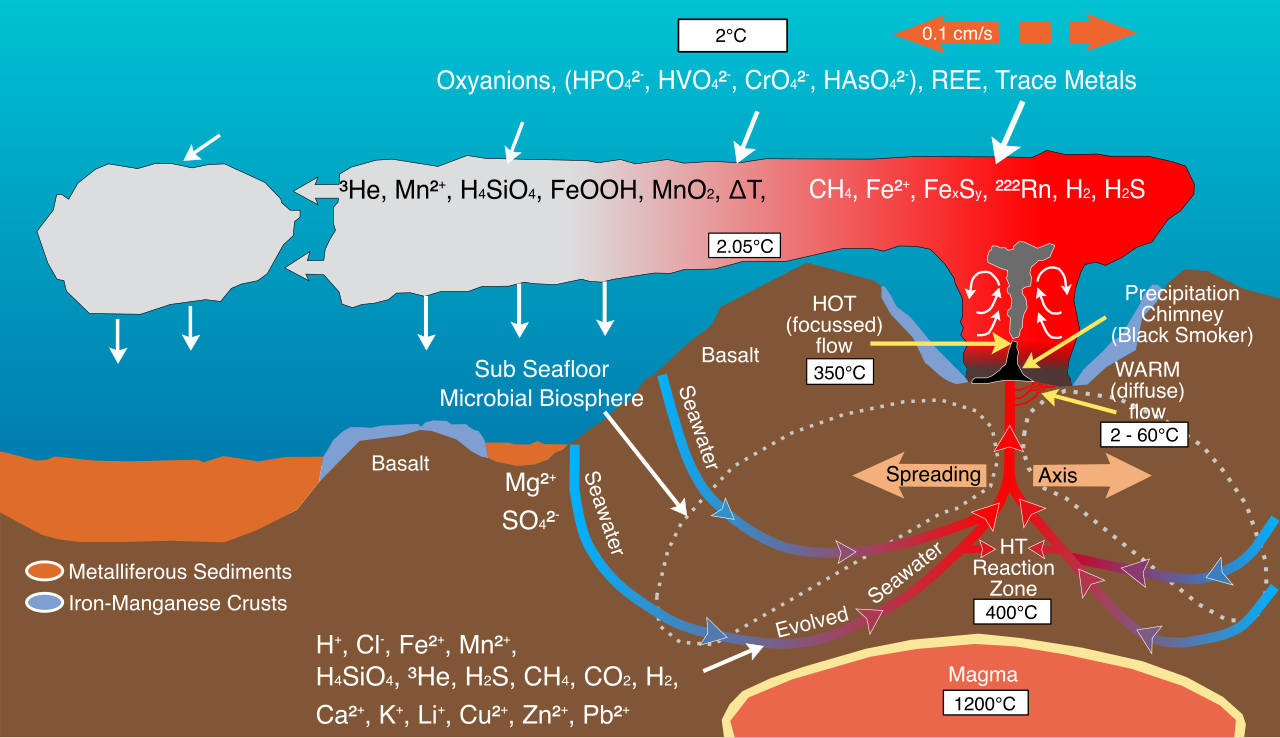
However, from an evolutionary and biochemical standpoint, photosynthesis and chemosynthesis are sibling processes. Here is how you can help students map one onto the other:
| Feature | Photosynthesis | Chemosynthesis |
|---|---|---|
| Energy Source | Photons (Sunlight) | Oxidation of inorganic chemicals |
| Carbon Source | Both photosynthesis and chemosynthesis fix inorganic carbon dioxide into organic sugar molecules. | Same |
| ATP Generation | Both photosynthesis and chemosynthesis utilize an electron transport chain to generate a proton gradient for ATP synthesis. | Same |
Final Thoughts for the Educator
When you explain photosynthesis, do not let your students get completely lost in the alphabet soup of RuBP, G3P, and 3-PGA without connecting it back to the grand picture. The thylakoid uses light to split water, charge batteries, and create an electrical gradient. The stroma uses those batteries to weld carbon gas into physical structure. Whether fueled by the blazing sun in an oak leaf or the scalding hydrogen sulfide of a deep-sea vent, life is simply a masterful game of electron capture. Teach the mechanics, but never lose the magic.