Ecosystems and Energy Flow
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
A single photon of light travels 93 million miles through the vacuum of space, only to strike the chloroplast of a blade of grass. In that microscopic collision, physics becomes biology. This is the fundamental engine of life on Earth: energy flows through an ecosystem in a single, unidirectional path, while matter constantly cycles through the living and non-living components of an ecosystem. As an educator preparing secondary students for the rigors of biological science, you are not merely teaching them to memorize vocabulary words or draw arrows on a food web; you are handing them the accounting ledger of the biosphere. They need to see that an ecosystem operates under strict physical laws, where every joule of energy is taxed by thermodynamics, and every carbon atom is endlessly repurposed. Let us examine the physical theaters where this drama unfolds, the rules that govern the biological transaction of energy, and the systemic consequences when those systems are perturbed.
Before tracking the flow of energy, we must establish the boundaries of the ecosystems themselves. Biomes are large-scale environments defined by distinct climatic conditions and characterized by adapted communities of organisms.
Terrestrial Biomes
Temperature and precipitation dictate the distribution of terrestrial biomes. Your students must be able to link the climatic drivers of a biome to the evolutionary adaptations of the organisms that live there.
| Biome | Geographic & Climatic Drivers | Distinguishing Characteristics |
|---|---|---|
| Tropical Rainforests | Located in equatorial regions; characterized by high annual rainfall and constant high temperatures. | Because of the ideal growing conditions, tropical rainforests possess the highest net primary productivity of any terrestrial biome. |
| Savannas | Tropical regions. | These are grasslands with scattered trees. Savannas experience distinct dry and wet seasons, forcing flora and fauna to adapt to temporary droughts. |
| Subtropical Deserts | Centered at approximately 30 degrees north and south latitude. | They experience very low and highly unpredictable precipitation. Consequently, plants in subtropical deserts exhibit physiological and structural adaptations to minimize water loss (e.g., deep taproots, waxy cuticles). |
| Chaparral | Coastal regions (like California or the Mediterranean). | The chaparral biome is characterized by dry summers and wet winters. Ecologically, the chaparral biome is dominated by shrubs adapted to periodic fires. |
| Temperate Grasslands | Mid-latitudes (like the American prairies). | Temperate grasslands experience pronounced annual fluctuations in temperature. Notably, trees are largely absent in temperate grasslands due to low precipitation and frequent fires. |
| Temperate Forests | Mid-latitudes with moderate rainfall. | Temperate forests experience defined growing seasons during the spring, summer, and early fall. They are primarily dominated by deciduous trees that drop leaves to conserve resources in winter. |
| Boreal Forest (Taiga) | Subarctic regions just south of the Arctic Circle. | The boreal forest is characterized by cold, dry winters and short, cool, wet summers. Rather than broadleaf trees, coniferous trees dominate the flora of the boreal forest biome. |
| Arctic Tundra | High northern latitudes. | The Arctic tundra is characterized by an extremely low average winter temperature. The defining soil feature is that the Arctic tundra contains a layer of permanently frozen soil called permafrost, which severely limits root depth. |
Aquatic Biomes
Aquatic ecosystems are categorized not by temperature and rainfall, but by depth, sunlight penetration, and water flow.
Oceanic Realms and Zones
- The pelagic realm comprises all of the open water in the ocean. Think of it as the vast, watery water column itself.
- The benthic realm encompasses the ocean bottom from the shoreline to the deepest parts of the ocean floor. It is the physical substrate.
- Moving vertically from the surface downward, the photic zone is the portion of the ocean that allows sufficient sunlight penetration for photosynthesis.
- Beneath it lies the aphotic zone, which is the ocean layer where sunlight cannot penetrate.
- The abyssal zone is the deepest part of the ocean and completely lacks sunlight, making it incredibly cold and highly pressurized.

Coastal and Continental Regions
- The intertidal zone is the oceanic region that is closest to land. Because of the tides, organisms in the intertidal zone must endure alternating periods of being submerged in water and exposed to air.
- Moving outward, the neritic zone extends from the intertidal zone to the edge of the continental shelf. This well-oxygenated, sunlight-rich zone is highly productive.
- Within some of these warm shallow waters, you will find coral reefs, which are ocean ridges formed by marine invertebrates.
Freshwater and Estuarine Systems
- Estuaries are biomes that occur where a source of fresh water meets the ocean. Because of the tidal wash, estuaries feature a mixing of fresh water and salt water, creating uniquely brackish habitats that serve as vital nurseries for marine life.
- Lakes and ponds are standing bodies of fresh water that can undergo thermal stratification during the summer months, leading to distinct temperature layers that isolate nutrients and oxygen.
- Rivers and streams are continuously moving bodies of fresh water, carrying nutrients from terrestrial headwaters down to lakes or oceans.
- Finally, wetlands are environments where the soil is permanently or periodically saturated with water, distinct from lakes because they feature emergent vegetation rooted in the soil but rising above the surface.
Life requires energy to combat entropy. This energy enters ecosystems via autotrophs.
- Photoautotrophs use solar energy to synthesize organic compounds. They are the gateway for sunlight to enter the biological world.
- Chemoautotrophs use inorganic molecules as an energy source to synthesize organic compounds. They are the foundation of ecosystems completely devoid of sunlight, such as hydrothermal vents in the abyssal zone.
When teaching productivity, give your students a financial analogy. Gross primary productivity (GPP) is the total rate at which photosynthetic producers incorporate energy from the sun. This is an ecosystem's "gross paycheck" before biological taxes. However, the plants themselves must survive; they burn some of that energy through cellular respiration. What is left over is net primary productivity (NPP)—the energy that remains in producers after subtracting the energy used for respiration.
Why does this distinction matter? Because net primary productivity represents the total amount of energy available to primary consumers. It is the actual, tangible budget upon which the rest of the ecosystem relies.
Once energy is converted into organic matter, it must be distributed.
- A food chain is a linear sequence of organisms through which nutrients and energy pass.
- Each organism in a food chain occupies a specific trophic level.
- Because nature is infinitely complex, a single chain rarely tells the whole story. A food web is a graphic representation of intersecting and interconnected food chains within an ecosystem.
Let's look at the specific trophic levels:
- Primary consumers are herbivores that consume primary producers.
- Secondary consumers are typically carnivores that consume primary consumers.
- Tertiary consumers are carnivores that consume secondary consumers.
- Apex consumers occupy the highest trophic level in a food web. They have no natural predators.
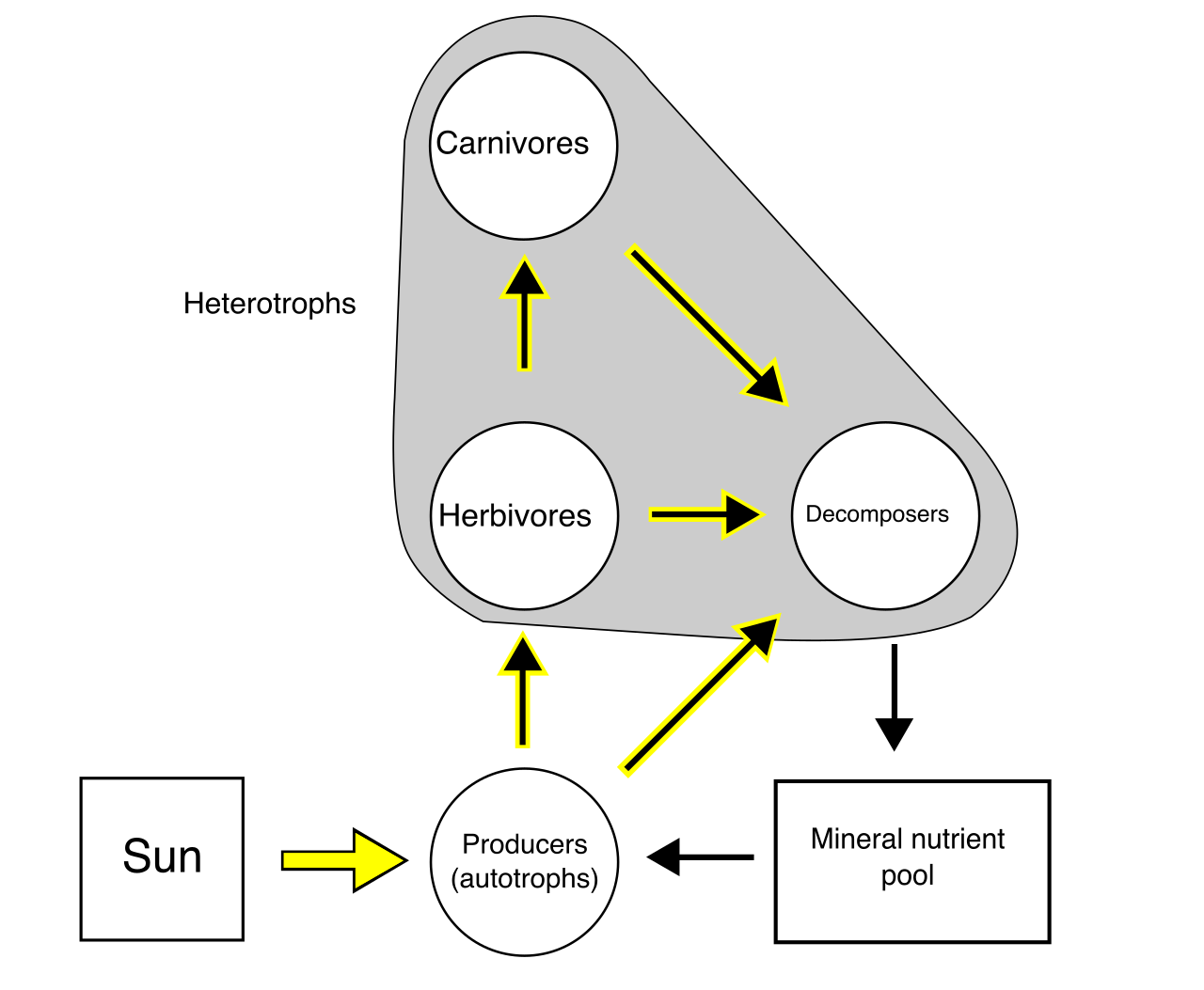
Teacher Tip: The Clean-Up Crew Distinction High school students frequently conflate detritivores and decomposers. You must draw a sharp mechanical distinction: Detritivores are organisms that ingest and internally digest dead organic matter (such as earthworms chewing through leaf litter). Conversely, decomposers secrete enzymes to externally break down dead organic material (such as fungi releasing enzymes into a fallen log, then absorbing the dissolved nutrients).
Why are there millions of blades of grass, thousands of gazelles, but only a handful of cheetahs on a savanna? The answer lies in ecological efficiency, which is the measurement of energy transfer efficiency between two successive trophic levels.
Due to the Second Law of Thermodynamics, energy conversions are terribly inefficient. In biological systems, approximately ten percent of the energy from one trophic level is successfully transferred to the next higher trophic level. The other 90% is lost to the environment as metabolic heat or trapped in indigestible waste. This mathematical reality has profound implications: The rapid loss of energy between trophic levels naturally restricts the maximum length of food chains. Most chains cap out at four or five links; there simply isn't enough thermodynamic budget to support apex predators above that.
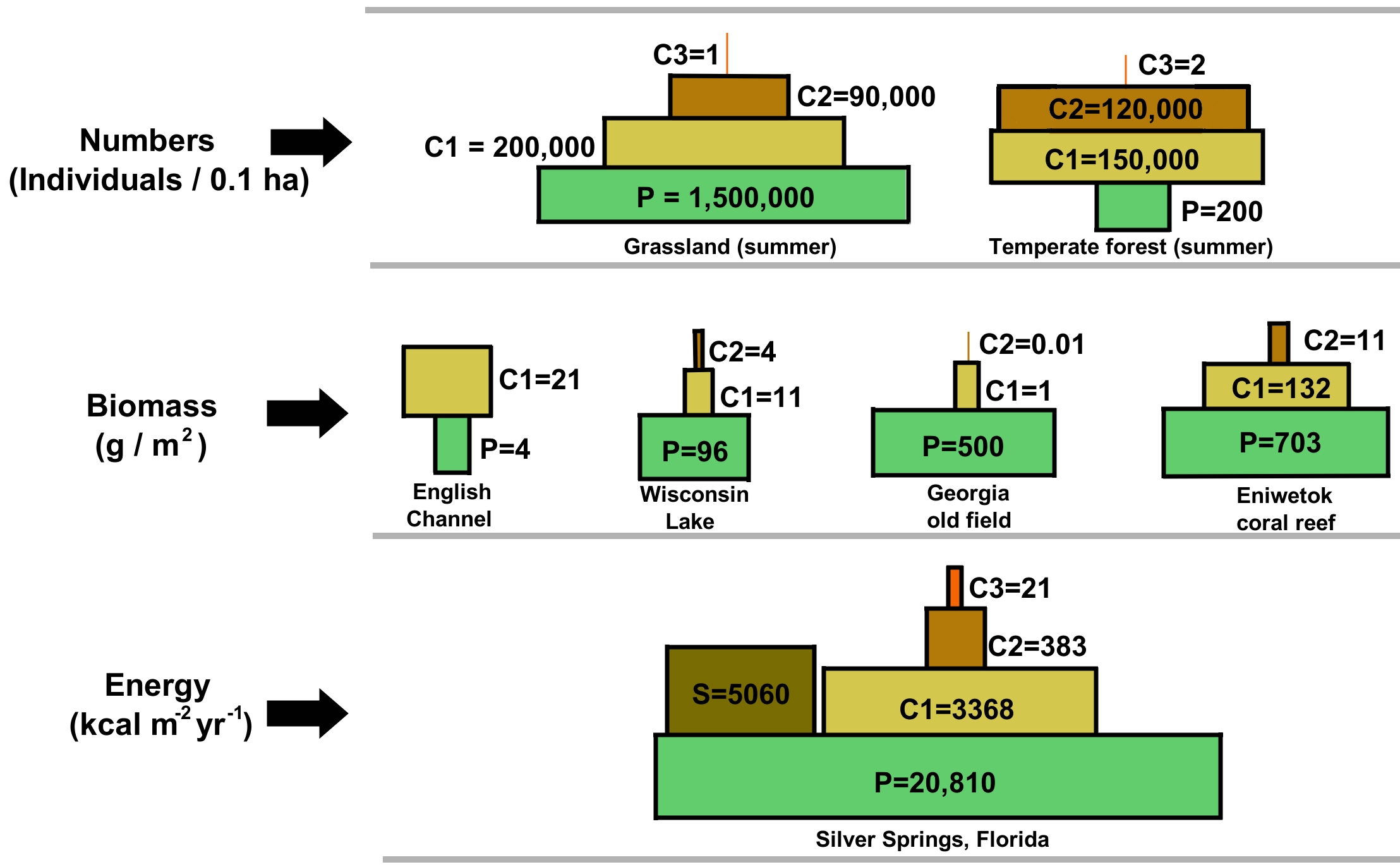
We visualize these realities using an ecological pyramid, which is a graphical representation used to quantify the trophic structure of an ecosystem.
1. Pyramids of Energy
Pyramids of energy represent the rate of energy flow through each successive trophic level. Because energy is constantly lost at each step, pyramids of energy are always upright. They can never be inverted due to the laws of thermodynamics.
2. Pyramids of Biomass
Pyramids of biomass represent the total dry mass of all living organisms at each trophic level at a specific time. (We use dry mass because water weight fluctuates and contains no usable metabolic energy).
While usually upright in terrestrial ecosystems, pyramids of biomass can be inverted in certain aquatic ecosystems. How is this geometrically possible without breaking the laws of physics? An inverted biomass pyramid occurs when primary producers have a very high reproduction and consumption rate. For example, in the ocean, the rapid turnover rate of aquatic phytoplankton allows a small standing biomass to support a larger standing biomass of zooplankton. The phytoplankton are eaten as fast as they reproduce, so their standing mass at any one snapshot in time is small, even though their productivity (energy flow) is enormous.
3. Pyramids of Numbers
Pyramids of numbers represent the exact count of individual organisms present at each trophic level. Like biomass pyramids, a pyramid of numbers can be inverted when a single large primary producer supports many small primary consumers. Think of a single massive oak tree (one producer) supporting a hundred thousand herbivorous caterpillars (100,000 primary consumers).
Because food webs are interconnected networks, pulling on one thread distorts the whole fabric. A trophic cascade occurs when the addition or removal of top predators causes reciprocal changes in lower trophic levels.
When an apex predator is removed, the population of their immediate prey (secondary or primary consumers) explodes. This overpopulation subsequently overgrazes the producers. Thus, a top-down trophic cascade often alters the abundance or biomass of primary producers in a food web, even though the predator never ate the plants directly.
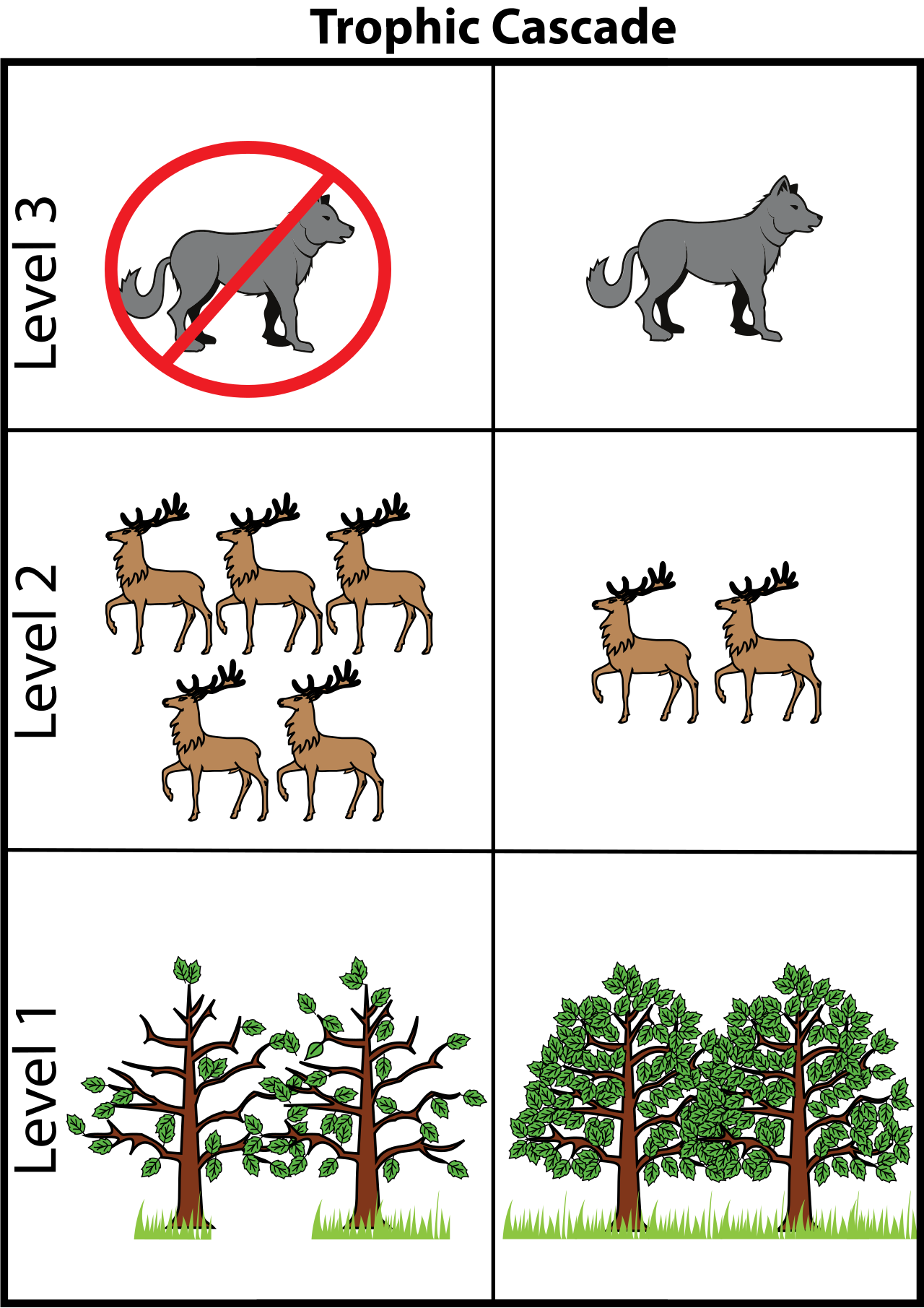
The ultimate textbook example you will teach is how the reintroduction of wolves to Yellowstone National Park triggered a well-documented trophic cascade. By preying on elk and changing their foraging behavior, the wolves allowed overgrazed willow and aspen along the riverbanks to recover, which in turn stabilized the riverbanks and brought back the beaver and songbird populations.
We have established that energy dissipates, but matter cycles. This creates a terrifying dynamic when human-made toxins enter a food web. As an educator, you must ensure your students understand the mechanistic difference between bioaccumulation and biomagnification.
Bioaccumulation occurs when a single organism absorbs a toxic substance at a rate faster than the substance is excreted. This happens within an individual's lifetime.
However, biomagnification is the increasing concentration of persistent toxic substances in organisms at successive higher trophic levels. This happens across the food chain.
For a chemical to threaten an ecosystem via biomagnification, it must meet specific chemical criteria:
- Substances that undergo biomagnification must be resistant to environmental degradation. If they break down quickly, they cannot travel up the chain.
- Chemicals that biomagnify are typically highly soluble in lipids. Because water-soluble toxins can be flushed out in urine, they generally do not biomagnify. Conversely, fat-soluble toxins are stored in the adipose tissue of organisms instead of being excreted.
When a pelican eats a hundred contaminated fish, it absorbs and forever stores the accumulated toxins of all one hundred fish into its own body fat. The higher the trophic level, the more concentrated the poison.
Real-world applications of this process are highly testable:
- Dichlorodiphenyltrichloroethane (DDT) is an insecticide historically known to cause severe biomagnification in bird populations. It disrupted calcium metabolism, causing apex birds like bald eagles to lay eggs with shells so thin they crushed under the weight of the incubating mother.
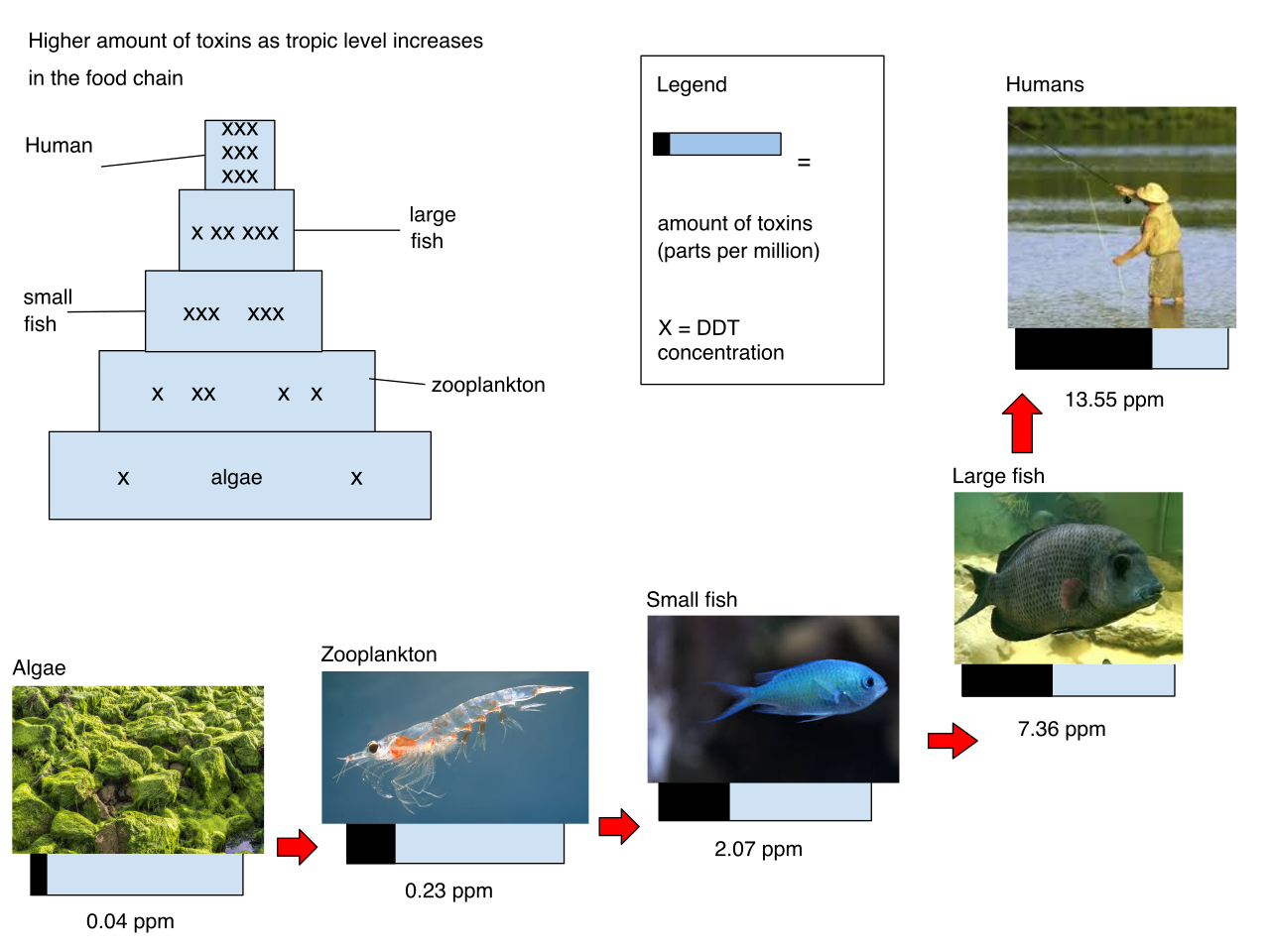
- Polychlorinated biphenyls (PCBs) are synthetic compounds that biomagnify significantly in aquatic food webs, building up in predatory fish and marine mammals.
- Heavy metals such as mercury and lead undergo biomagnification in aquatic and terrestrial ecosystems. This is exactly why pregnant women are advised to avoid eating apex predatory fish like swordfish or king mackerel.
When you stand before your students, remember that you are explaining the fundamental operating system of the natural world. Energy drives the engine; matter builds the parts. By mastering these principles, your students will look at a forest, a pond, or a savanna and see the profound biological accounting taking place right beneath their eyes.