Non-Mendelian Inheritance
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
[Gregor Mendel]]'s [monastery garden offered a beautifully sanitized view of inheritance. By carefully selecting traits in pea plants that sorted independently and exhibited strict dominant-recessive hierarchies, Mendel established the foundational laws of genetics. But nature is rarely so polite. Non-Mendelian inheritance encompasses the vast array of genetic patterns that deviate from the phenotypic ratios predicted by Gregor Mendel's laws. Biological reality is wonderfully messy. Genes interact, mask one another, travel together on chromosomes, and even respond directly to the temperature of the air. For an aspiring biology educator, this is where genetics comes alive. When a student asks why they are taller than both their parents, or why tortoiseshell cats are almost exclusively female, Mendelian laws fall short. We must look to the spaces between, where inheritance reveals itself not as a simple coin flip, but as a complex, dynamic system.
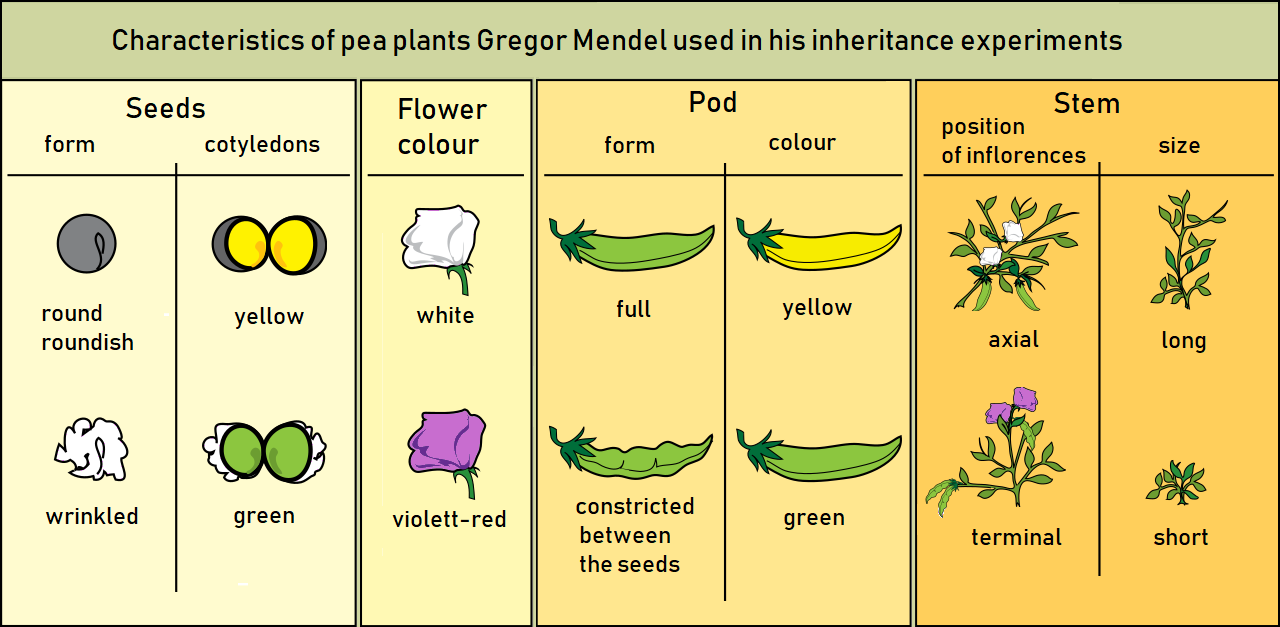
Mendel taught us that one allele dominates another completely. If we view inheritance through this lens alone, we miss the nuances of how proteins actually interact in a cell.
Incomplete Dominance and Codominance
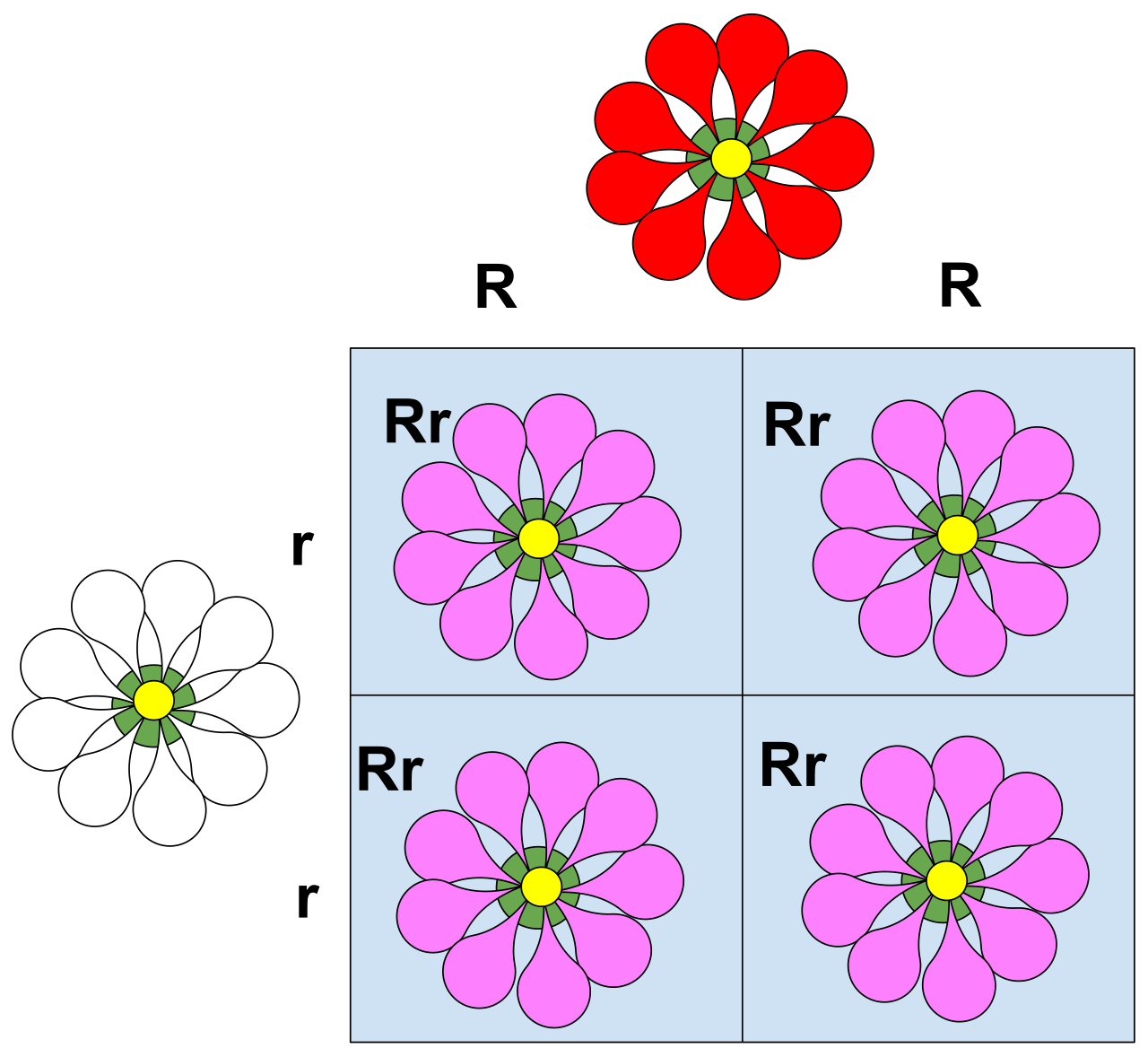
Sometimes, alleles do not conquer; they compromise. Incomplete dominance produces a heterozygous phenotype that is an intermediate blend of both homozygous parental phenotypes. A classic classroom example is the pink flower color of heterozygous snapdragons resulting from a cross between red and white parents. The red allele does not produce enough pigment to fully mask the lack of pigment from the white allele, leaving us with pink.
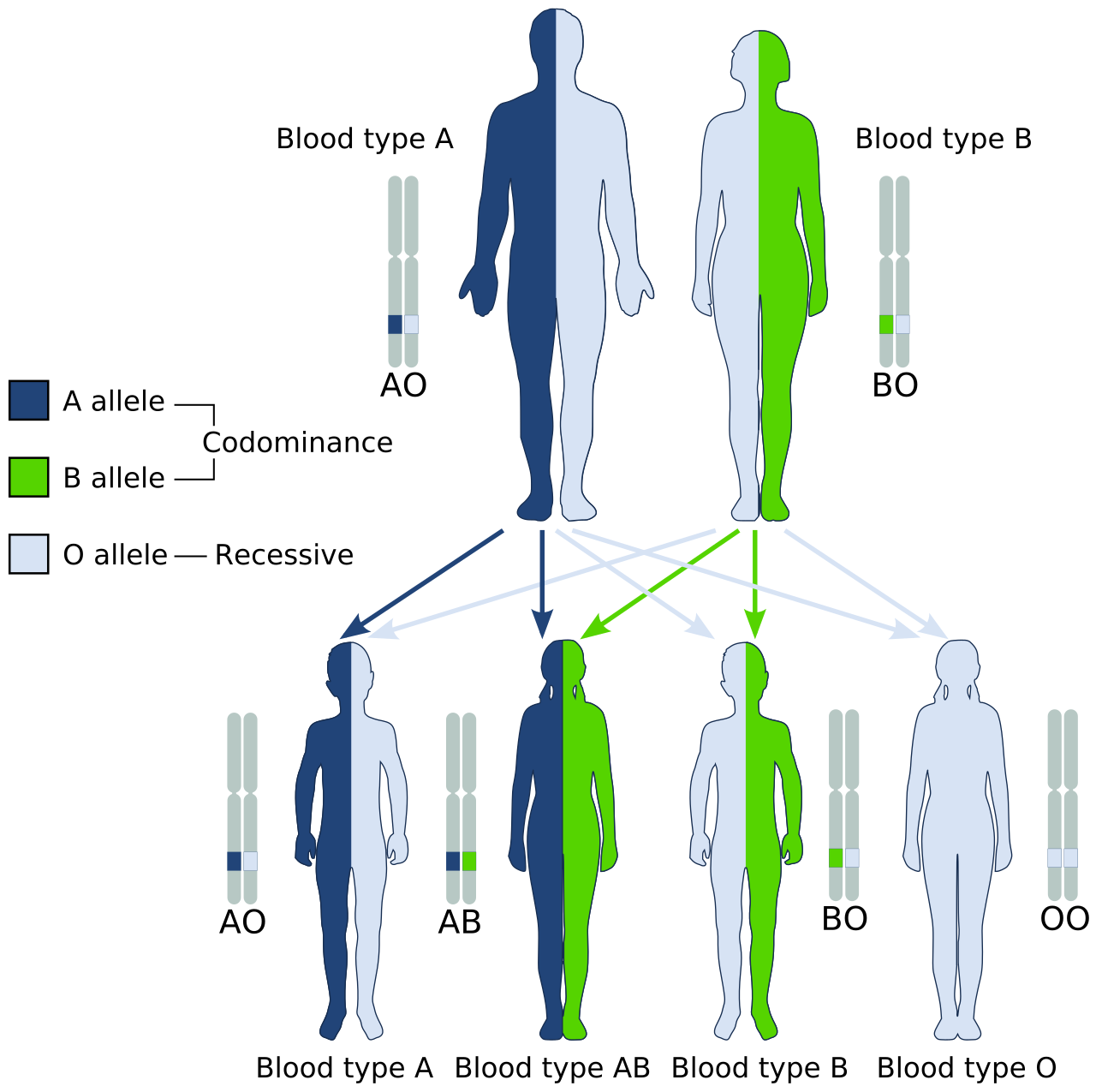
Codominance, by contrast, is not a blend but a collaboration. It results in the full and simultaneous expression of both alleles in a heterozygous individual. Consider human blood types. The simultaneous expression of A and B antigens in human blood type AB is an example of codominance. Both glycoproteins are proudly displayed on the surface of the red blood cell.
Multiple Alleles
In Mendel's world, genes came in two flavors: tall or short, green or yellow. In reality, a gene is a sequence of DNA subject to mutation, meaning multiple alleles exist when a population contains more than two allelic variants for a single gene locus. However, the mechanics of meiosis demand a strict physical limit: a single diploid organism possesses only two alleles for a specific gene locus regardless of the population's total allele count. You only have two slots—one on your maternal chromosome and one on your paternal chromosome—no matter how many variants exist in the human species.
To formalize how we discuss variants in a population, geneticists use the wild-type designation, which describes the most common phenotype or genotype found in a natural population. Any genetic variant that deviates from the wild-type sequence is termed a mutant allele.
Traits rarely exist in a vacuum. The most common misconceptions your future students will have stem from the idea of "one gene equals one trait."
| Concept | Mechanism | Real-World Example |
|---|---|---|
| Pleiotropy | A genetic condition involving a single gene that influences multiple distinct phenotypic traits. | Sickle cell anemia is a pleiotropic disorder causing red blood cell deformation along with multiple secondary health complications (e.g., organ failure, joint pain). |
| Polygenic Inheritance | Occurs when multiple distinct genes interact to determine a single phenotypic trait. | Human skin color and adult human height are examples of polygenic traits. |
| Epistasis | Occurs when the expression of one gene completely masks the phenotypic expression of a different gene. | The gene controlling pigment deposition in Labrador retrievers is epistatic to the gene controlling the specific pigment color (black vs. brown). |
Notice the mathematical symmetry here. Pleiotropy is a one-to-many relationship. Polygenic inheritance is a many-to-one relationship, which is why polygenic traits display a continuous variation of phenotypes within a population (a bell curve of heights, rather than just "tall" and "short"). Epistasis is a hierarchical relationship—if a Labrador retriever inherits two recessive alleles for pigment deposition, the dog will be yellow regardless of whether its color genes code for black or chocolate.
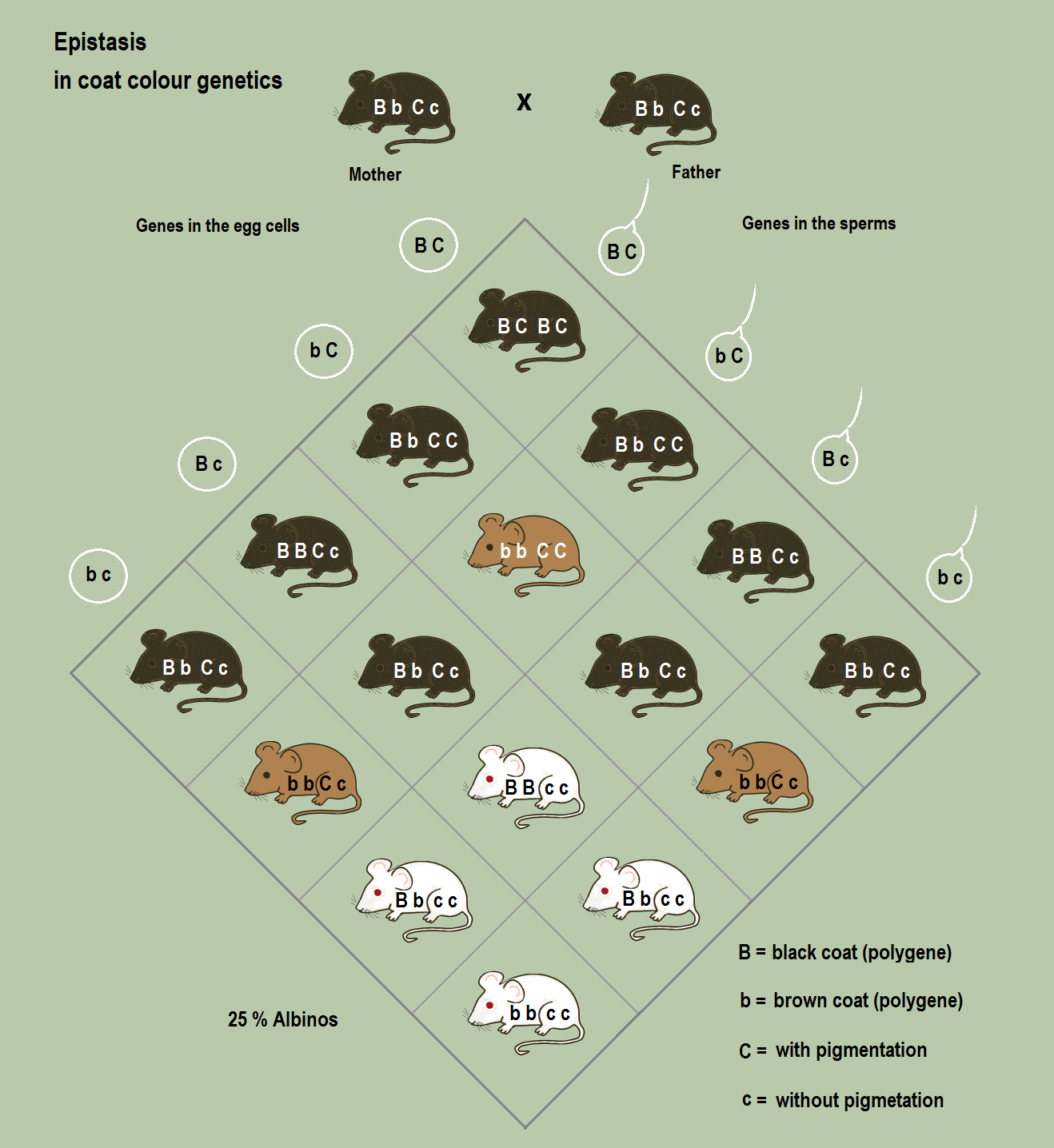
Sometimes, the mere presence of certain alleles alters the survivability of the organism.
- Recessive lethal alleles cause the death of an organism only when present in a homozygous state. For example, the tailless trait in Manx cats results from a recessive lethal allele that causes embryonic death when homozygous. Two tailless Manx cats will yield a 2:1 ratio of living offspring (tailless to normal), fundamentally breaking Mendel's expected 3:1 monohybrid ratio because the homozygous recessive embryos never survive to be counted.
- Dominant lethal alleles cause the death of an organism in both homozygous and heterozygous states. Because these alleles kill their host, they are rarely passed on, unless the lethality strikes after reproductive age. Huntington's disease is an example of a dominant lethal allele in humans that evades natural selection by remaining asymptomatic until mid-life.
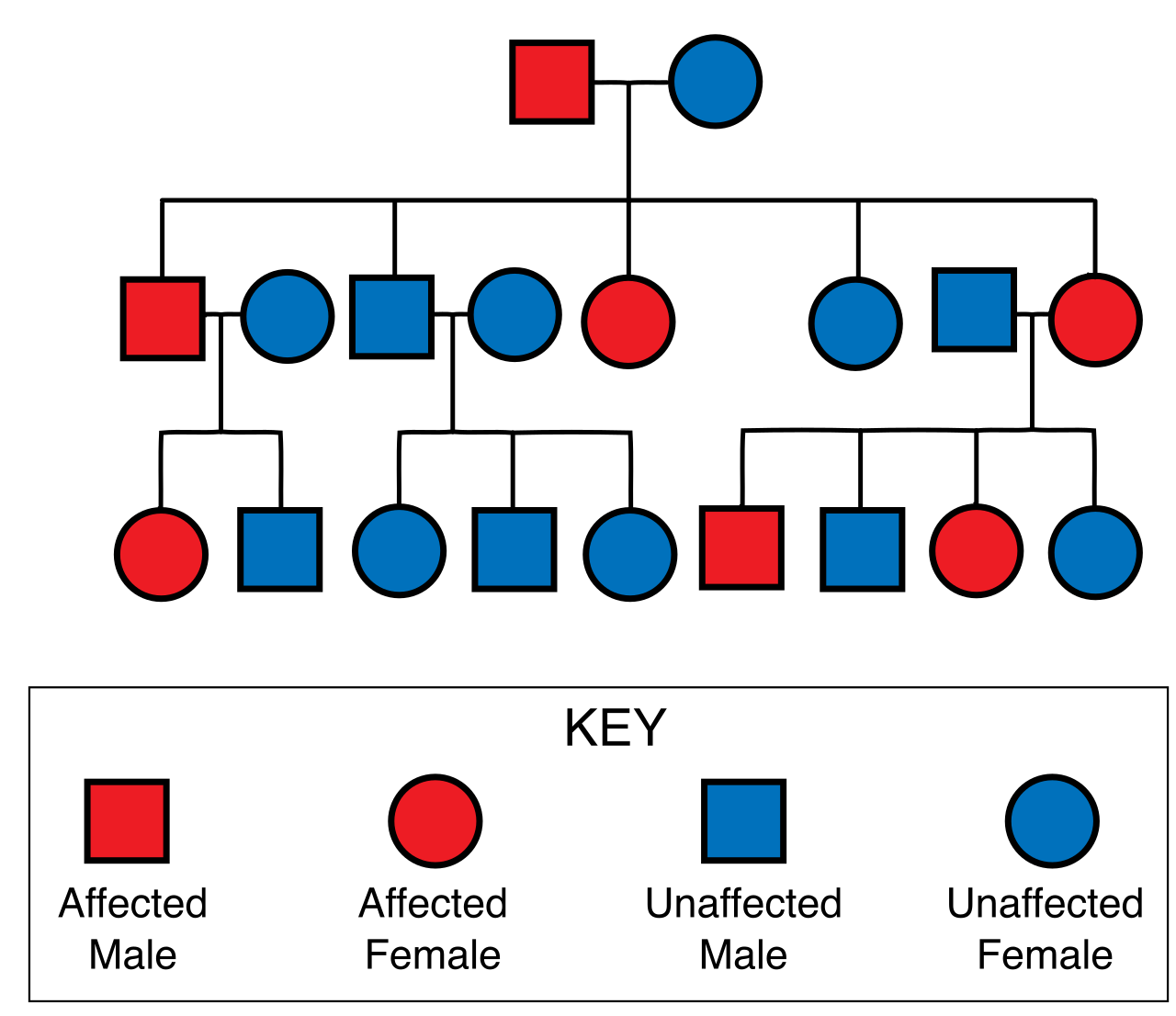
Even when an organism possesses a specific genotype, the physical outcome is not always a guarantee.
- Penetrance defines the percentage of individuals with a specific genotype who actually express the expected phenotype. (Think of this as binary: did the trait appear across the population, yes or no?)
- Expressivity defines the degree of phenotypic intensity expressed by a particular genotype in an individual organism. (Think of this as a volume knob: how loud is the trait in this specific individual?)
Mendel’s Law of Independent Assortment works perfectly when genes reside on different chromosomes. But humans have over 20,000 genes packed onto just 23 pairs of chromosomes. Gene linkage violates Mendel's law of independent assortment because linked genes are physically located close to each other on the same chromosome.
Because of this physical proximity, linked genes are frequently inherited together as a single unit during meiosis. They refuse to sort independently.
However, the linkage is not unbreakable. Homologous recombination (crossing over) during prophase I of meiosis can separate linked genes onto different chromatids. The physical architecture of chromosomes provides us with an elegant mathematical reality:
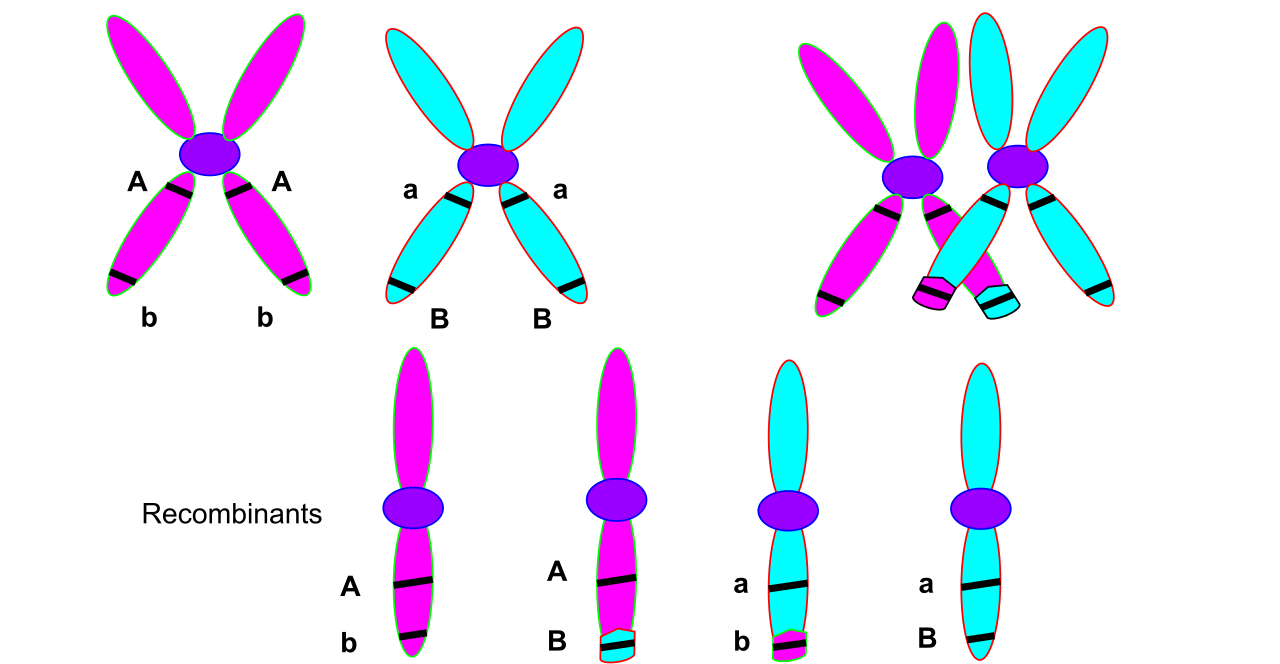
The recombination frequency between two genes is directly proportional to their physical distance on a chromosome. One map unit or centimorgan represents a one percent recombination frequency between two linked genes.
By observing how often traits are inherited together versus how often they cross over, you can physically map the chromosome. There is a ceiling to this metric: recombination frequencies cannot exceed fifty percent for any two genes. If genes are far enough apart, crossing over becomes a statistical certainty. Consequently, genes with a fifty percent recombination frequency assort independently despite being on the same chromosome.
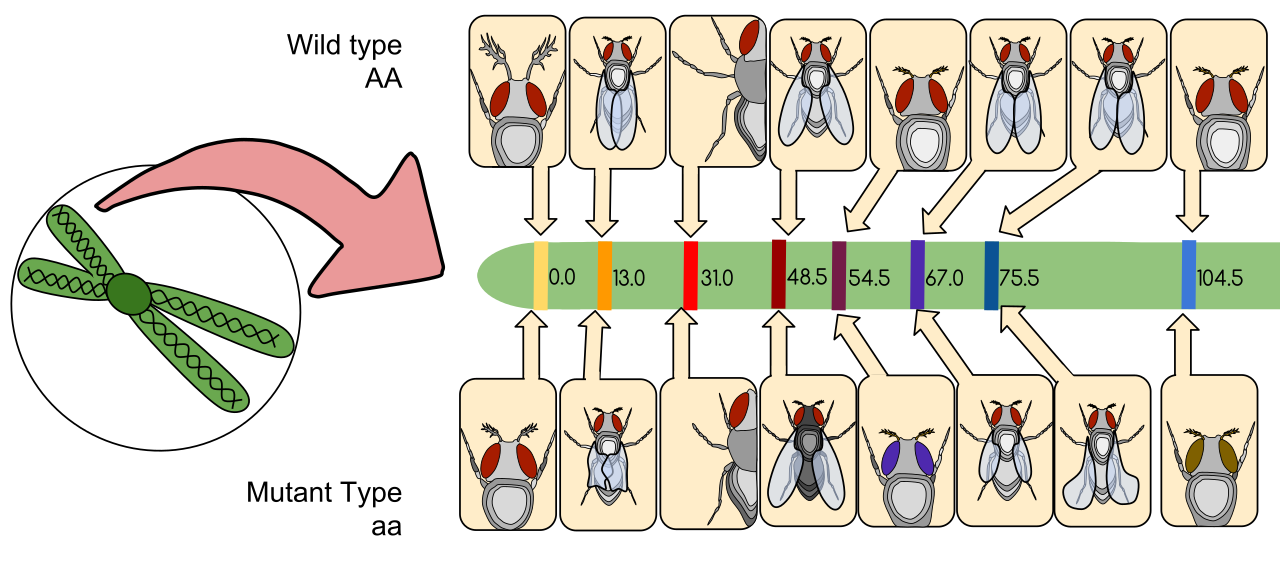
The discovery that chromosomes actually dictate traits was famously solidified by Thomas Hunt Morgan, who discovered sex-linked inheritance through his research on eye color in Drosophila melanogaster (the fruit fly).
Sex-linked inheritance involves genes located on the sex chromosomes rather than autosomes. Because the X chromosome is massive compared to the diminutive Y chromosome, most sex-linked traits are X-linked. This creates an imbalance. Human males are hemizygous for X-linked genes due to possessing only a single X chromosome. If a male inherits a recessive allele on his lone X chromosome, he expresses it—there is no second X to mask it.
Consequently, X-linked recessive genetic disorders appear more frequently in males than in females. A female must inherit two copies of a recessive X-linked allele to express the associated recessive trait, whereas a male needs only one.
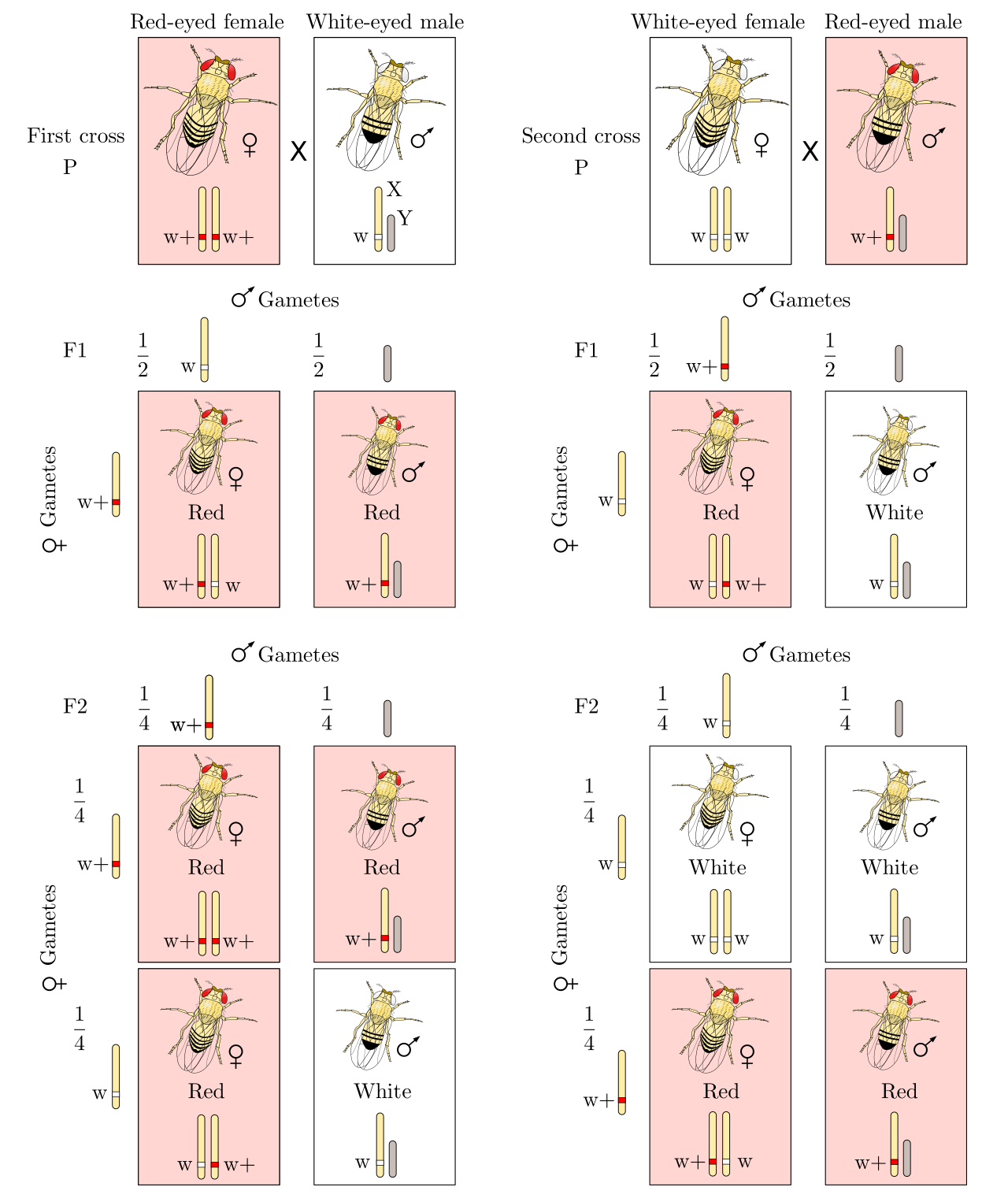
X-Inactivation
If females have two X chromosomes and males have one, females produce twice the amount of X-linked proteins, right? No. Mammalian evolution solved this dosage problem with an elegant mechanism: X-inactivation.
X-inactivation randomly condenses one X chromosome into an inert Barr body in female mammals. This crucial biological process equalizes the dosage of X-linked gene products between mammalian males and females.
Because this inactivation happens randomly in each cell during early embryonic development, females are genetic mosaics. A striking, visible result of random X-inactivation is the tortoiseshell coat color in female cats. In one patch of skin, the X chromosome carrying the orange fur allele is active; in a neighboring patch, the X chromosome carrying the black fur allele is active.

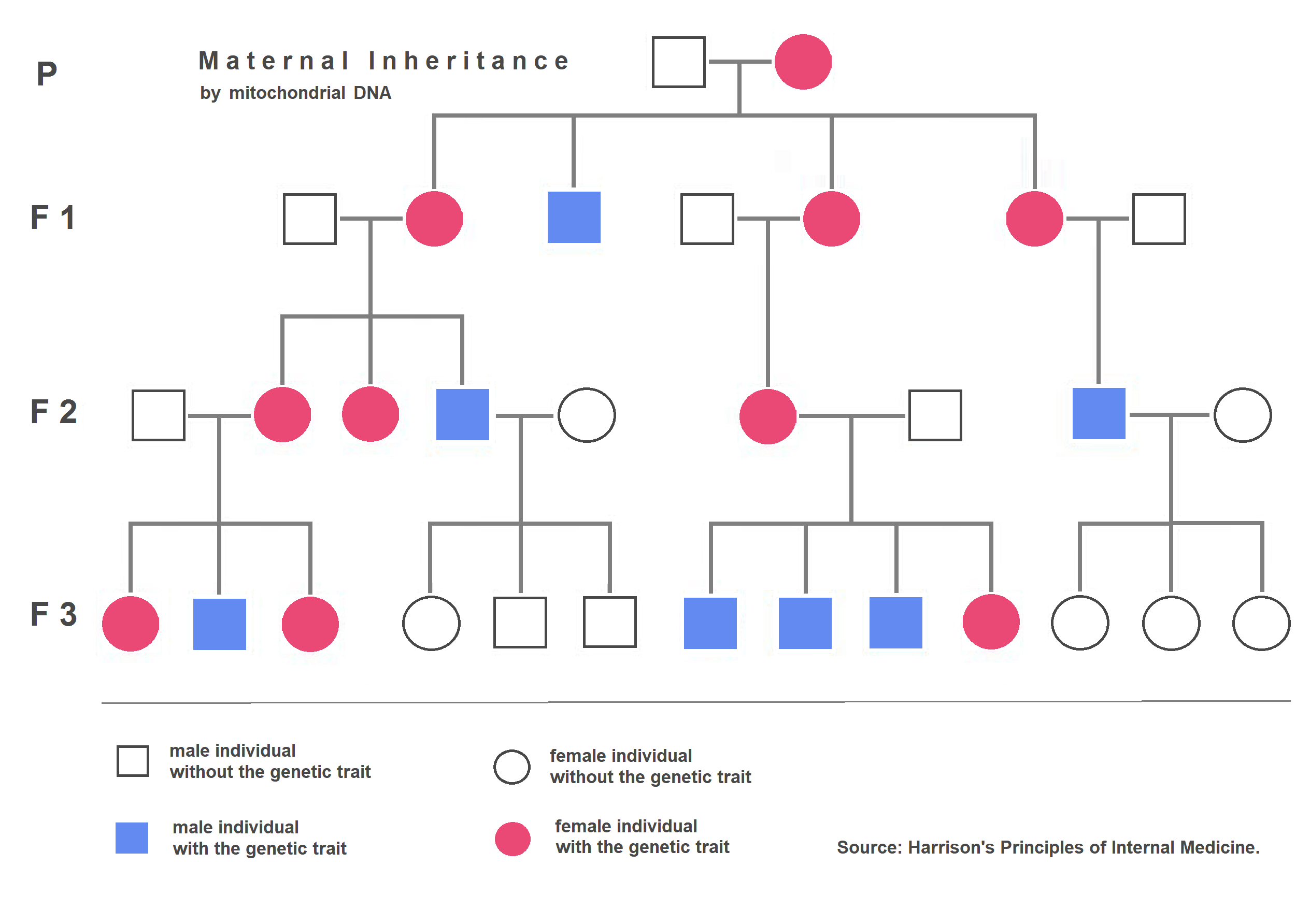
We instinctively visualize genetic material as safely locked inside the nucleus. Yet, extranuclear inheritance refers to the transmission of genes located in organelles outside the cellular nucleus.
Mitochondria and chloroplasts contain independent circular DNA molecules, a biochemical echo of their ancient origins as free-living bacteria. During fertilization, the massive egg provides essentially all the cytoplasm—and therefore all the organelles—to the zygote. Consequently, human mitochondrial DNA is inherited exclusively from the maternal parent.
This brings us to a related phenomenon known as a maternal effect, which occurs when the phenotype of the offspring is determined solely by the genotype of the mother. Even if the offspring carries a dominant paternal allele, the maternal proteins deposited into the egg long before fertilization dictate the early developmental trajectory of the zygote.
Genetics is not destiny; it is a blueprint handed to the environment. Environmental factors can directly alter the phenotypic expression of a specific genotype.
- Temperature: Temperature-sensitive alleles govern the localized coat color expression in Siamese cats and Himalayan rabbits. The enzyme producing melanin degrades at core body temperatures, meaning pigment is only deposited in the cooler extremities of the animal (ears, nose, paws, tail).

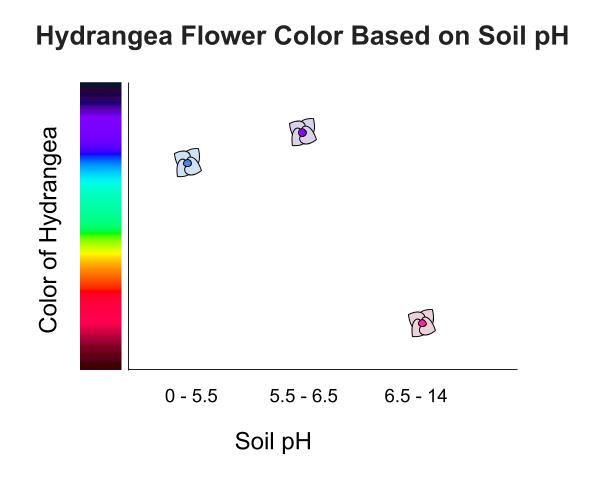
- Chemistry: Soil pH determines the flower color of Hydrangea plants by altering aluminum availability in the ground. Acidic soils yield blue flowers, while alkaline soils yield pink ones.
Epigenetics
Finally, we arrive at the frontier of modern genetics: Epigenetics. This involves heritable changes in gene expression without any alteration to the underlying DNA nucleotide sequence. It is how cells with identical DNA—like a neuron and a liver cell—look and function entirely differently. They "read" different parts of the genetic library based on chemical tags.
Two primary mechanisms drive this:
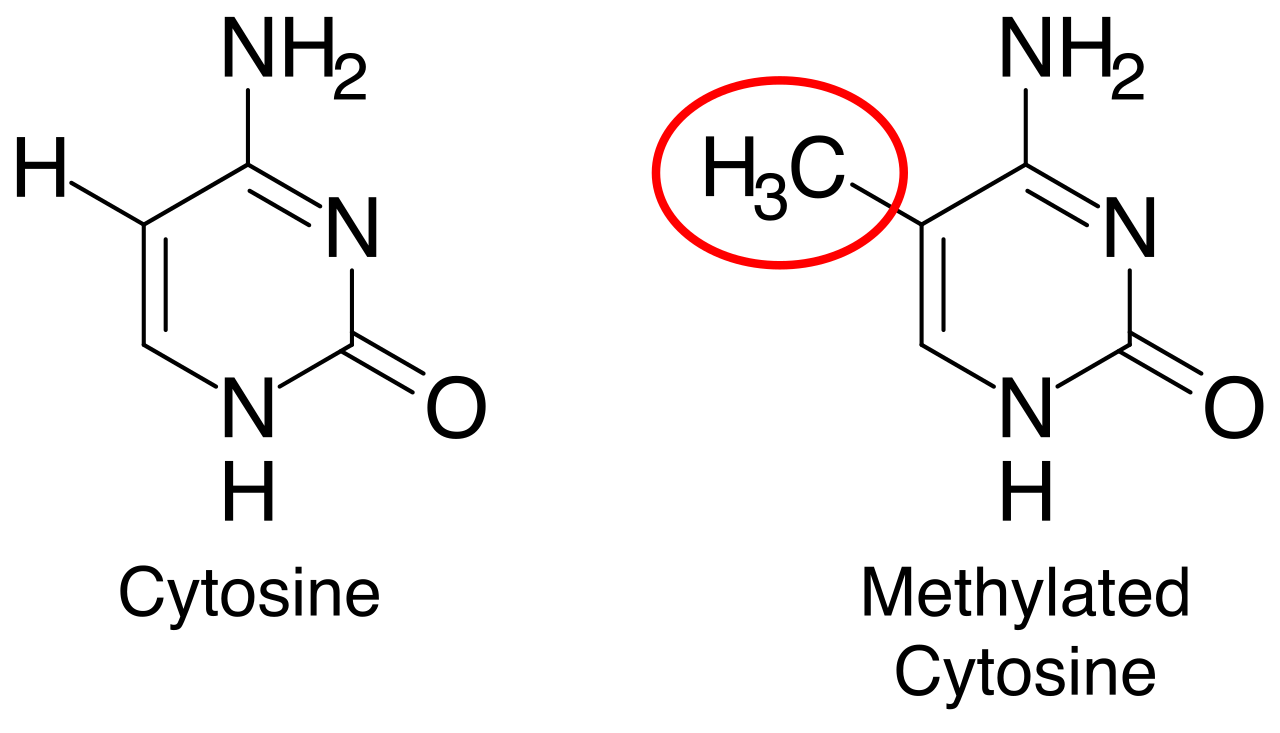
- DNA methylation: The addition of methyl groups physically obstructs transcription factors to repress gene transcription. (It acts as a molecular "off switch.")
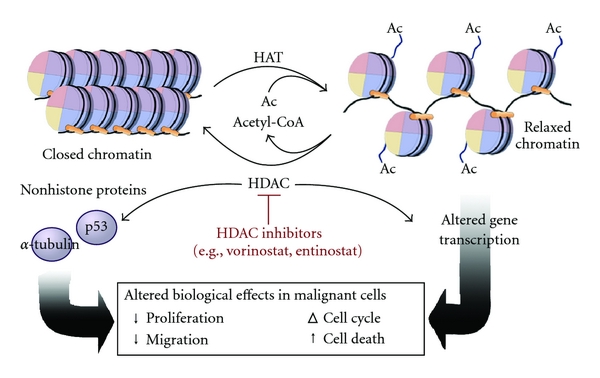
- Histone acetylation: The addition of acetyl groups to histone proteins loosens the association between DNA and histones to enhance gene expression. (It acts as an "on switch," unwinding the DNA so it can be read.)
These epigenetic markers can even be passed down depending on which parent supplied the chromosome. Genomic imprinting is an epigenetic phenomenon causing gene expression to depend on the parent of origin. For certain imprinted genes, the maternal copy is actively methylated and silenced, while only the paternal copy is expressed (or vice versa).
As a biology educator, your objective is to guide students from the clean mathematics of Punnett squares to the beautiful, chaotic reality of molecular interactions. By mastering non-Mendelian inheritance, you prepare them to see biology not as a static set of rules, but as an elegant, adaptable system of life.