Population Genetics and Speciation
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
A population is not a static portrait; it is a statistical engine constantly shuffling a massive, dynamic pool of alleles. Every new generation represents a new statistical sample, subject both to the random walks of probability and the rigid, deterministic sieve of the environment. To master population genetics is to understand exactly how the frequency of these alleles changes over time—and how those shifting frequencies eventually fracture a single lineage into the vast, branching diversity of life.
As a future biology teacher, you will encounter a persistent misconception: your students will inherently view evolution as a conscious, goal-oriented march toward perfection, driven entirely by natural selection. Your task is to dismantle this teleological illusion. You must show them that evolution is a tapestry woven from multiple, distinct threads—some deterministic, some entirely random, and some driven by geographic accident.
Here is the definitive guide to the mechanisms that alter allele frequencies, the evolutionary patterns they produce, and the precise mechanical barriers that create new species.
Evolution at its most fundamental level is simply a change in allele frequencies within a population over time. While natural selection acts on fitness, other mechanisms drive genetic change purely through migration, human intervention, or sheer mathematical randomness.
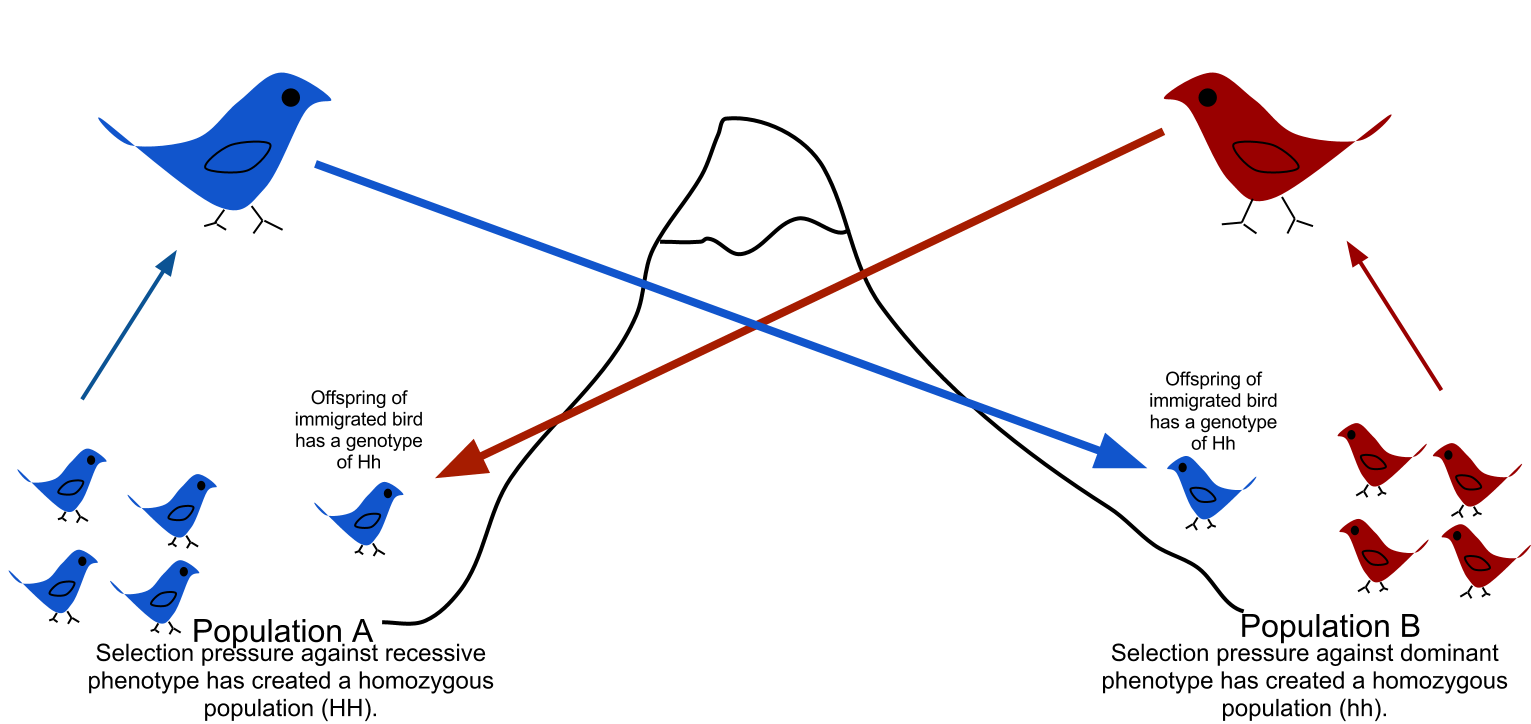
Gene Flow: The Great Homogenizer
Imagine two isolated lakes, each containing a distinct population of fish. If a flood temporarily connects these lakes, fish from one lake swim into the other. This is gene flow, which is the transfer of genetic variation from one population to another through the migration of individuals or gametes (such as pollen blowing across a valley).
Gene flow has two immediate mathematical consequences. First, gene flow introduces new alleles into a recipient population, which fundamentally increases genetic variation within a receiving population. However, when viewed on a macro level across both populations, high rates of gene flow reduce genetic differences between distinct populations over time. If individuals constantly migrate back and forth, the two distinct pools blend into a single, genetically uniform ocean.
Genetic Drift: The Blind Hand of Probability
While gene flow is about movement, genetic drift is about sampling error. Genetic drift is the change in the frequency of an existing gene variant in a population due to random sampling of organisms. Unlike natural selection, which cares about how well a trait helps an organism survive, genetic drift is completely blind to biological fitness.
The Mathematics of Drift: The effects of genetic drift are mathematically stronger in small populations compared to large populations. If you flip a coin 10 times, getting 8 heads (80%) is not unimaginable. If you flip it 1,000 times, getting 800 heads is practically impossible.
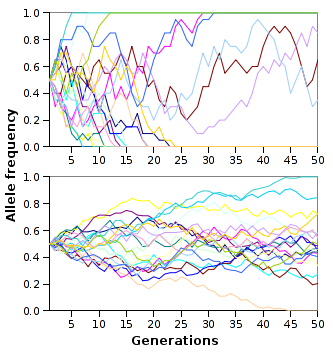
In small populations, this random sampling can act with ruthless finality. Over successive generations, genetic drift can lead to the complete loss of a specific allele from a population, wiping out genetic diversity regardless of whether that allele was beneficial. Conversely, genetic drift can cause a specific allele to become fixed in a population at a frequency of 100 percent, meaning every future individual will inherit it unless a new mutation arises.
Drift manifests most dramatically in two distinct scenarios:
-
The Bottleneck Effect: This occurs when a population's size is drastically reduced for at least one generation, usually by a catastrophic event like a fire or disease. The surviving individuals are not necessarily the most fit; they are simply the luckiest. Consequently, the bottleneck effect results in a newly formed population with a different allele frequency than the original population.
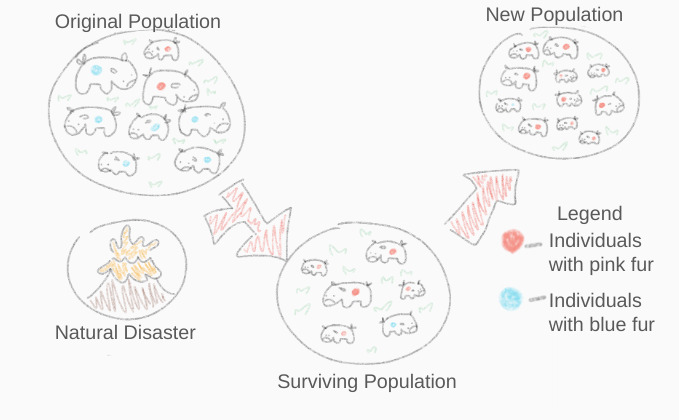
The bottleneck effect demonstrates how a catastrophic reduction in population size leaves a random assortment of survivors, drastically altering the allele frequencies of the new population. Source: Bottleneck Effect by Ihatezxmbyz, CC BY-SA 4.0. -
The Founder Effect: This occurs when a small number of individuals break off from a larger population to establish a new colony. Because the "founders" carry only a tiny fraction of the original population's genetic diversity, the founder effect generally results in reduced genetic variation in the new colony compared to the source population.
Sexual and Artificial Selection: The Biased Sieve
Natural selection is a struggle for survival, but survival means nothing if an organism fails to reproduce.
Sexual selection is a specific mode of natural selection where members of one biological sex choose mates of the other sex to mate with. This evolutionary pressure often defies basic survival logic—a peacock's massive, brilliantly colored tail makes it highly visible to predators, but without it, the male will never attract a mate. Furthermore, sexual selection includes competition between members of the same sex for access to members of the opposite sex (e.g., male elk locking antlers).

Because these reproductive pressures affect males and females differently, sexual selection frequently leads to sexual dimorphism, which is the systematic difference in form between individuals of different sex in the same species.
Conversely, when humans step into the role of the environment, we get artificial selection, which is the identification by humans of desirable traits in plants and animals. In artificial selection, humans direct the mating process to enhance and perpetuate desirable traits in future generations. Every dog breed, from the Great Dane to the Chihuahua, and every crop in our modern agricultural system is a testament to the speed and power of artificial selection.

As these microevolutionary forces operate over millions of years, they trace distinct geometric patterns across the tree of life. When teaching these patterns, anchor them directly to the anatomical structures they produce.
| Pattern | Definition & Mechanism | Structural Outcome & Significance |
|---|---|---|
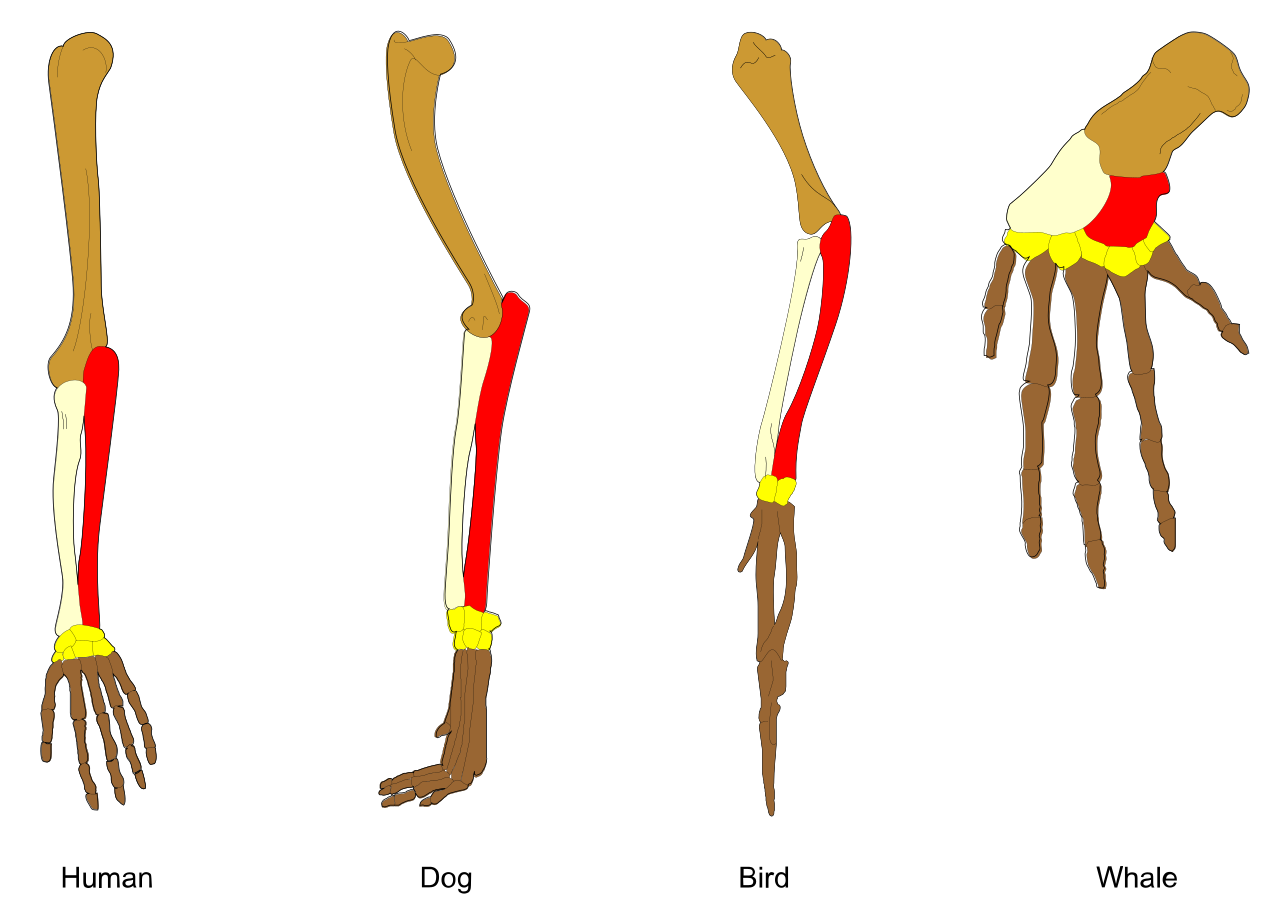
| Divergent Evolution | The accumulation of differences between closely related populations. | Divergent evolution produces homologous structures that share a similar embryonic origin (e.g., the human arm and the whale flipper). Over time, divergent evolution typically leads to speciation. |
| Convergent Evolution | The independent evolution of similar features in species of different evolutionary lineages. | Convergent evolution creates analogous structures that have similar form or function but were not present in the last common ancestor (e.g., the wings of a bat and the wings of an insect). |
| Coevolution | Occurs when two or more species reciprocally affect each other's evolutionary trajectory. | The evolutionary arms race between a predator and prey is an example of coevolution (e.g., a toxic newt and the garter snake evolving resistance to the toxin). |
| Adaptive Radiation | A process in which organisms diversify rapidly from an ancestral species into a multitude of new forms. | Adaptive radiation commonly occurs when a change in the environment opens new ecological niches (e.g., the extinction of the dinosaurs allowing early mammals to rapidly diversify). |
We have established how populations change, but at what point does a changed population become an entirely new entity?
Speciation is the evolutionary process by which populations evolve to become distinct species. But defining exactly what a "species" is requires precision. In population genetics, we rely heavily on the biological species concept, which defines a species as a group of organisms that can successfully interbreed and produce fertile offspring.
If two organisms cannot produce fertile offspring, they are, by this definition, distinct species. The mechanisms that enforce this boundary are known collectively as reproductive isolation.
Reproductive isolation is a collection of evolutionary mechanisms, behaviors, and physiological processes critical for speciation. Ultimately, the purpose of reproductive isolation prevents members of different species from producing viable, fertile offspring.
We categorize these barriers based on when they act: before or after the formation of a zygote.
Prezygotic Barriers
Prezygotic reproductive barriers prevent mating or fertilization between different species. They are nature's frontline defense against hybridization.
- Habitat isolation is a prezygotic barrier where two species live in the same area but occupy different habitats (e.g., one snake lives in the water, a closely related species lives in the trees; they never meet).
- Temporal isolation is a prezygotic barrier where species breed at different times of the day, seasons, or years. An orchid that blooms in April cannot cross-pollinate with an orchid that blooms in July.
- Behavioral isolation is a prezygotic barrier where species have unique courtship rituals that prevent mate recognition. If a female firefly does not recognize the specific blinking rhythm of a male, she will not mate with him.
- Mechanical isolation is a prezygotic barrier where morphological differences prevent successful mating. The literal "lock and key" of their reproductive organs do not fit.
- Gametic isolation is a prezygotic barrier where the sperm of one species is unable to fertilize the eggs of another species, often due to hostile chemical environments in the reproductive tract or incompatible protein receptors on the egg's surface.
Postzygotic Barriers
If a sperm successfully fertilizes an egg across species lines, postzygotic reproductive barriers occur after a hybrid zygote is formed, serving as a secondary fail-safe.
-
Reduced hybrid viability is a postzygotic barrier where hybrid embryos fail to develop or survive to maturity. The genetic instructions from the parents are simply too conflicting to build a functional organism.
-
Reduced hybrid fertility is a postzygotic barrier where hybrid offspring are vigorous but remain sterile. The classic example you should keep in your pedagogical toolkit: mules represent an example of reduced hybrid fertility because mules are sterile offspring of a male donkey and a female horse. The mule is robust and strong, but because horses have 64 chromosomes and donkeys have 62, the mule ends up with 63—an odd number that prevents normal meiosis.

Mules are the robust but sterile offspring of a male donkey and a female horse. Because their parents have mismatched chromosome counts, mules cannot undergo normal meiosis, creating a permanent postzygotic barrier. -
Hybrid breakdown is a postzygotic barrier where first-generation hybrids are viable and fertile but subsequent generations are feeble or sterile. The system works for one generation, but the genetic recombination in the second generation causes the genome to collapse.
Speciation relies on the reproductive barriers listed above, but how do populations arrive at these barriers in the first place? It usually begins with how they are physically situated in space.
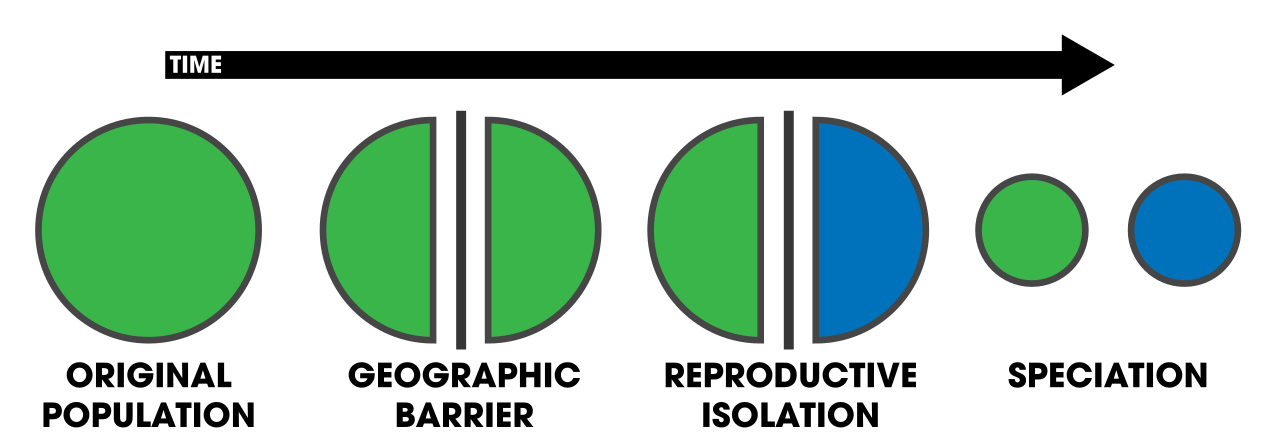
Allopatric Speciation: The Physical Divide
Allopatric speciation occurs when biological populations of the same species become geographically isolated from each other.
Imagine a river carving a deep canyon through a plateau over millennia, separating a population of ground squirrels into two disconnected groups. Geographic isolation in allopatric speciation prevents gene flow between the separated populations. Without gene flow to homogenize them, the two squirrel populations are now subjected independently to genetic drift, different mutations, and potentially different selective pressures on either side of the canyon. Over time, they diverge so deeply that if they were to meet again, prezygotic or postzygotic barriers would prevent them from interbreeding.
Sympatric Speciation: Division from Within
Sympatric speciation is the evolution of a new species from a surviving ancestral species while both continue to inhabit the same geographic region.
To your students, this will initially seem counterintuitive. If there is no physical barrier, why doesn't gene flow keep the population united? Sympatric speciation requires specialized mechanisms to spontaneously sever gene flow:
-
Polyploidy: This is a sudden, massive chromosomal mutation. Polyploidy is the condition of possessing more than two complete sets of chromosomes (e.g., a diploid plant producing a tetraploid offspring due to a failure in meiosis). Because a tetraploid cannot easily cross back with a diploid to produce fertile offspring, reproductive isolation is instantaneous. Consequently, polyploidy is a primary mechanism of sympatric speciation in flowering plants. Therefore, sympatric speciation can occur through polyploidy entirely within the span of a single generation.
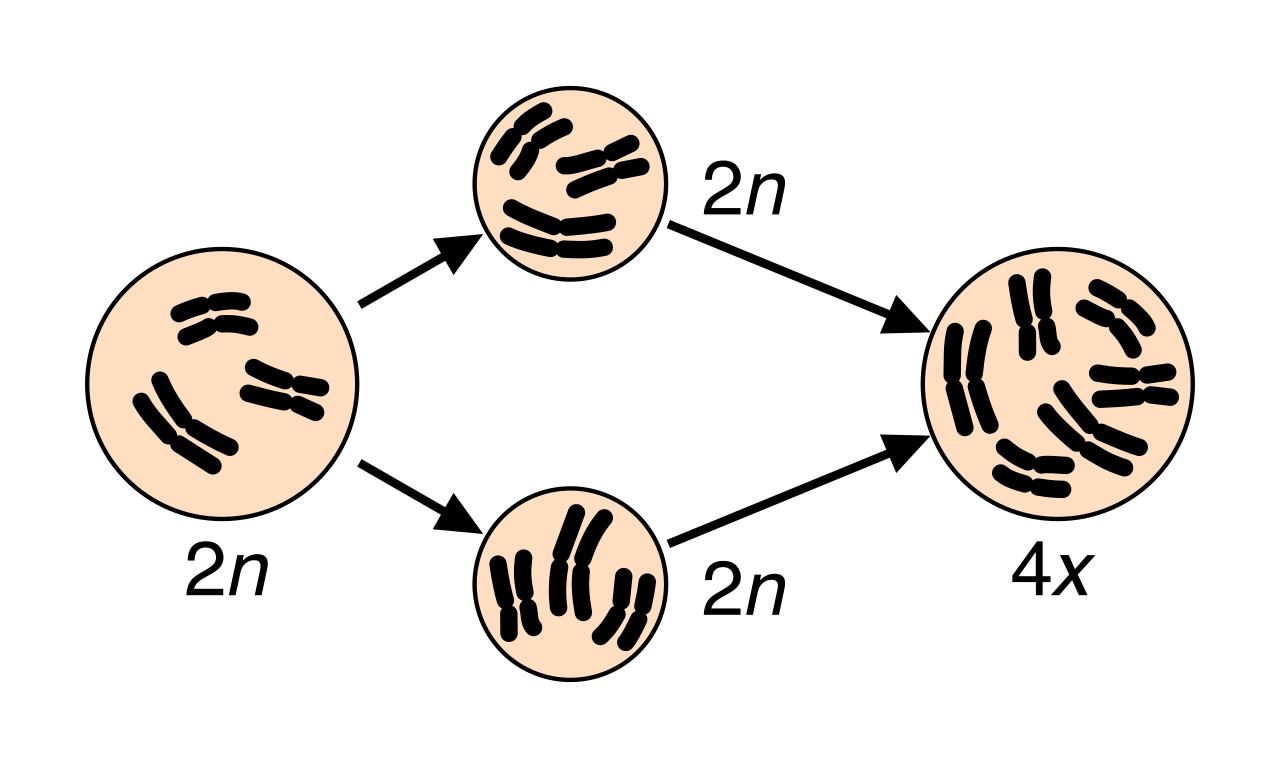
Speciation via polyploidy can occur instantly. A failure in meiosis produces diploid gametes, which self-fertilize to form a tetraploid zygote that is completely reproductively isolated from its diploid parents. -
Ecological Niche Partitioning: Sympatric speciation can occur when a subpopulation exploits a habitat or resource not used by the parent population. For example, if a subset of flies in a forest begins laying eggs on apples instead of the native hawthorn fruit, they will begin to interact and breed only with other apple-dwelling flies. Habitat isolation emerges without geographic separation.
-
Sexual Selection: Sympatric speciation can be driven by sexual selection when mate choice is based on distinct coloration or behaviors within the same habitat. In the hyper-diverse cichlid fishes of Lake Victoria, subtle shifts in the light-sensitivity of female eyes led them to prefer males of different colors. Because females only mated with specific color morphs, the single population fractured into hundreds of distinct, sympatric species sharing the exact same water column.
As an educator, your mastery of these concepts ensures you are not just teaching a vocabulary list, but revealing the grand, mechanical logic of life. By understanding the interplay between gene flow, genetic drift, selection, and the precise boundaries of reproductive isolation, your students will begin to see every species not as a static noun, but as an active, ongoing verb.