Evidence and Models of Evolution
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
To reconstruct the history of life on Earth is to read a massive, ancient ledger where many pages have been lost to fire and time, yet the overarching plot remains astonishingly clear. We do not need a time machine to verify evolutionary theory; the evidence is physically etched into the crust of the Earth, written into the anatomical blueprints of developing embryos, and coded directly into the chemical structure of every living cell. As an educator preparing to teach secondary biology, your objective is not to hand students a static timeline. Instead, you are teaching them how to read these geological and molecular archives—how to look at a whale's pelvis, a continent's coastline, or a sequence of DNA and deduce the invisible, billion-year history of life.
The fundamental starting point for understanding life's history is the earth itself. Fossils provide a visual record of extinct organisms in successive layers of rock, giving us tangible proof of organisms that no longer exist. When viewed as a whole, the fossil record demonstrates the chronological progression of life on Earth over billions of years. But to read this record, we must understand how to tell time geologically.
There are two primary ways scientists date fossils:
- Relative Dating: This method relies heavily on the law of superposition, which states that older sedimentary rock layers lie beneath younger sedimentary rock layers. By applying this law, relative dating allows us to estimate the comparative age of fossils simply by looking at their vertical placement in a cliff face or canyon.
- Radiometric Dating: When we need exact numbers, we turn to physics. Radiometric dating determines the absolute age of fossils using the decay rates of radioactive isotopes found in the surrounding rock. This relies on the concept of a half-life, which is the time required for exactly half of the radioactive isotopes in a given sample to decay.

As the timeline of life took shape, paleontologists discovered transitional fossils—extraordinary specimens that exhibit traits common to both an ancestral group and a derived descendant group. They are the "smoking guns" of evolutionary biology.
- Archaeopteryx is a classic transitional fossil showing features of both non-avian dinosaurs (teeth, long bony tail) and modern birds (flight feathers).
- Tiktaalik is another crucial transitional fossil, showing features of both lobe-finned fish (gills, scales) and early tetrapods (a functional neck, weight-bearing wrists), capturing the exact evolutionary moment vertebrates began to crawl onto land.

Life did not evolve on a static stage. Biogeography is the study of the geographic distribution of organisms on Earth, and it reveals that the global distribution of organisms is heavily influenced by the historical movement of tectonic plates.
Consider the breakup of the ancient supercontinent Pangaea. The breakup of Pangaea explains the presence of closely related fossils—like the ancient fern Glossopteris or the reptile Mesosaurus—on widely separated modern continents, such as South America and Africa. They didn't swim across the Atlantic; the land beneath them tore apart.
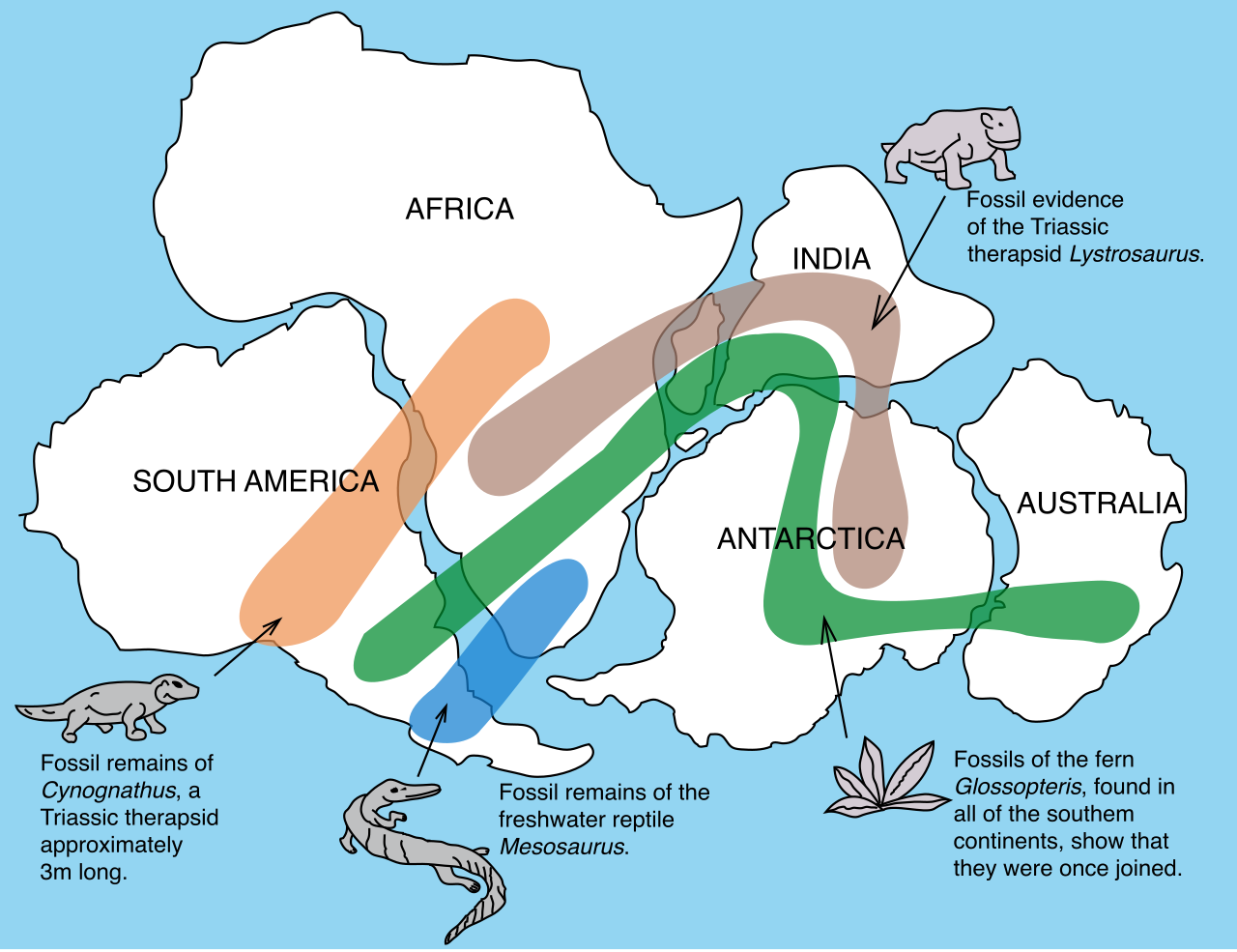
This geographic isolation drives speciation. We see this vividly in endemic species, which are species found naturally in only one specific geographic area.
- Oceanic Islands: These isolated landmasses often contain endemic species that are closely related to species on the nearest mainland (e.g., the Galápagos finches). The mainland provided the ancestral pioneers, while the island's isolation allowed them to evolve into entirely new forms.
- Continental Isolation: Marsupials diversified extensively in Australia due to prolonged geographic isolation from placental mammals. Without competition from placental mammals, marsupials radiated to fill nearly every ecological niche on the continent.
Evolution is a supreme recycler. It rarely builds from scratch; rather, it modifies existing structures to adapt to new environmental pressures.
Divergent Evolution and Homology
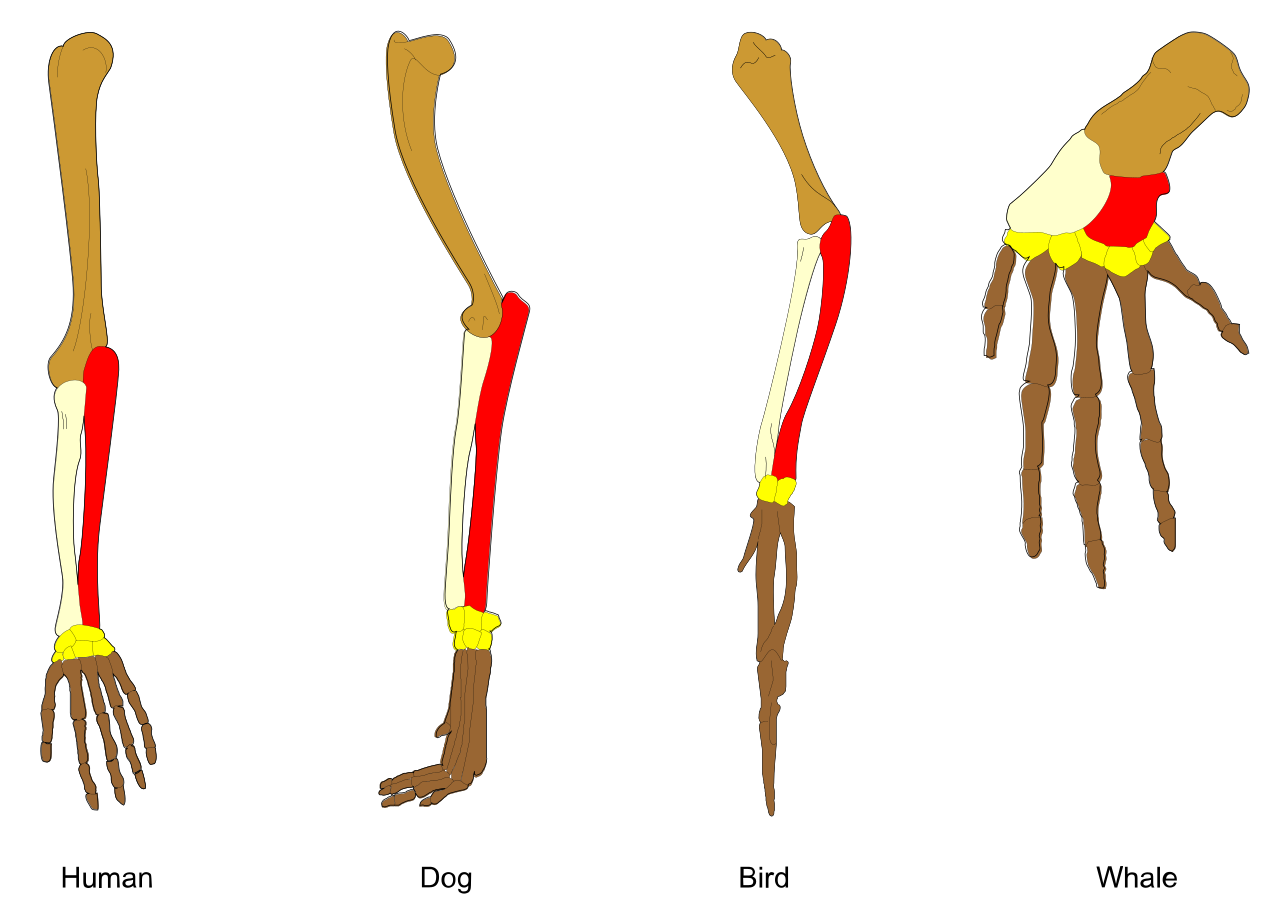
When two or more species evolve distinct traits from a common ancestor, we call it divergent evolution. This process produces homologous structures, which share a similar basic anatomy due to descent from a common ancestor. Because of divergent evolution, homologous structures frequently serve different functions in different modern species.
Example: Human arms, dog forelegs, bird wings, and whale flippers are homologous structures. They look different on the outside and perform different tasks (grasping, running, flying, swimming), but beneath the skin, they share the exact same skeletal arrangement of one thick bone, two thinner bones, and a cluster of wrist bones.
Convergent Evolution and Analogy
Conversely, convergent evolution occurs when unrelated organisms independently evolve similar traits due to similar environmental pressures. This results directly in analogous structures—traits that have similar functions without sharing a common evolutionary anatomical origin.
Example: The wings of insects and the wings of birds are analogous structures. Both solve the aerodynamic problem of flight, but their structural engineering is entirely different because they did not inherit wings from a common ancestor.
Vestigial Structures
Sometimes, a modified structure loses its utility entirely. Vestigial structures are anatomical features that have lost all or most of their original function in a species. They are the biological equivalents of silent letters in a word—remnants of traits that were fully functional in evolutionary ancestors.
- The human appendix is an example of a vestigial structure, a remnant of a larger cecum used by our ancestors to digest tough plant matter.
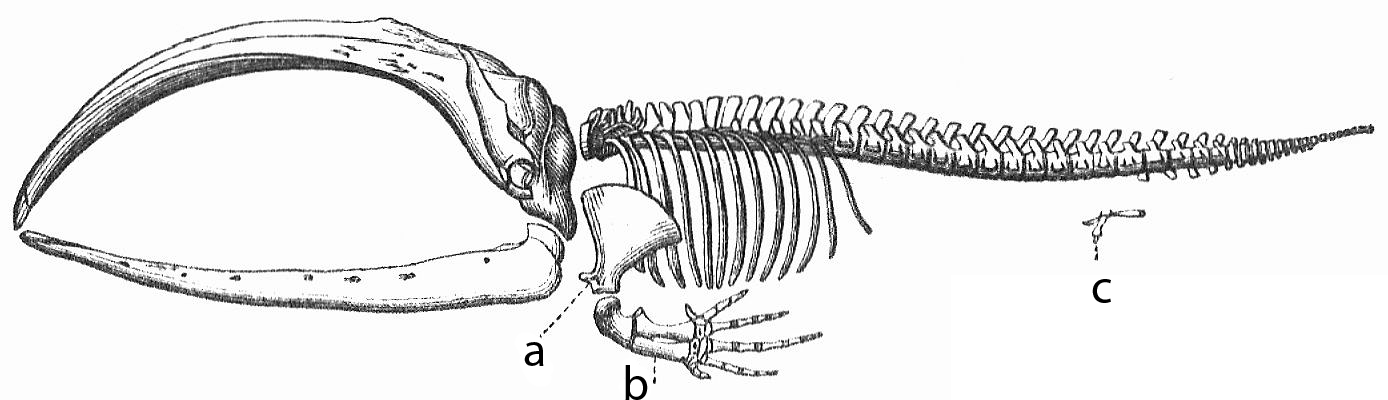
- The pelvic bones of modern whales are vestigial structures inherited from terrestrial ancestors. Whales have no hind legs, yet deep within their blubber, they carry a disconnected pelvis.
Embryology
Even when adult organisms look entirely different, their embryonic forms often betray their shared past. Shared embryonic structures provide evidence of common ancestry among morphologically diverse adult organisms. For instance, embryology reveals that all vertebrate embryos possess pharyngeal slits at some point during development. In fish, these become gills; in humans, they develop into parts of the jaw and the inner ear.
The most irrefutable evidence for evolution lies at the microscopic level. All living organisms use DNA as their primary genetic material. Furthermore, the genetic code for translating mRNA into proteins is nearly universal across all domains of life. Whether you are a bacterium, a mushroom, or a high school biology teacher, the codon AUG codes for the amino acid methionine. A universal genetic code strongly supports the hypothesis of a single common ancestor for all known life.
By sequencing genomes, we can quantify relatedness. Comparisons of DNA sequences reveal the degree of evolutionary relatedness between different species, with closely related species sharing a higher percentage of identical DNA sequences than distantly related species.
- Cytochrome c is a highly conserved protein used to study evolutionary relationships among aerobic organisms. Because it is essential for cellular respiration, mutations to it are rarely tolerated, making it a reliable marker for ancient evolutionary splits.
- Genomes are also littered with genetic fossils known as pseudogenes, which are non-functional DNA segments that structurally resemble functional genes. Shared pseudogenes between species indicate descent from a common ancestor that possessed the functional gene. If humans and chimpanzees both share the same deactivated gene for vitamin C synthesis, complete with the exact same disabling mutation, it is overwhelmingly probable they inherited that broken gene from a shared ancestor.
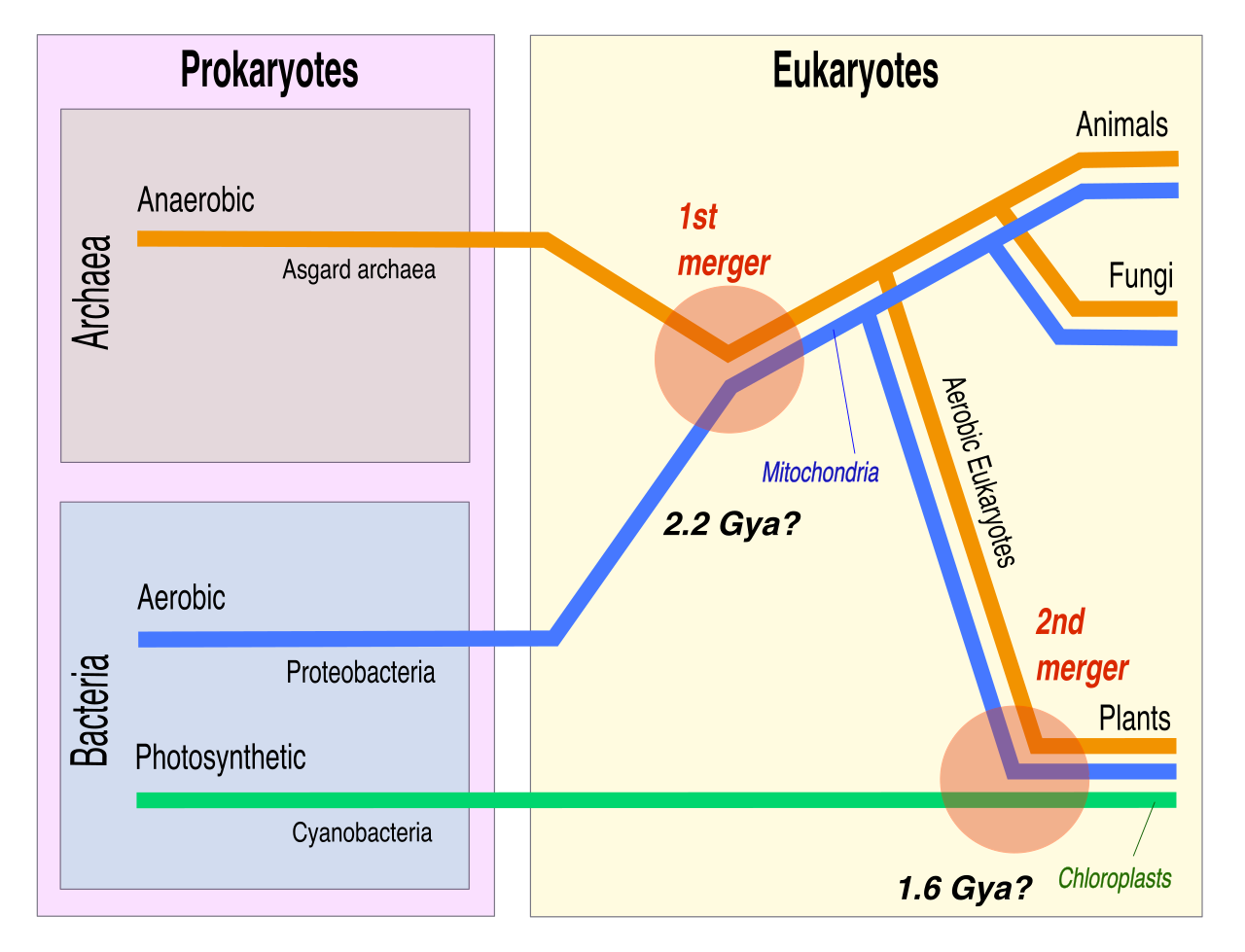
The Endosymbiotic Theory
One of the most profound molecular discoveries explains the very origin of complex eukaryotic cells. The endosymbiotic theory proposes that eukaryotic cells evolved from a symbiotic relationship involving engulfed prokaryotic cells.
Specifically:
- Mitochondria evolved from engulfed aerobic alpha-proteobacteria.
- Chloroplasts evolved from engulfed photosynthetic cyanobacteria.
The evidence for this is staggering and heavily tested. Both mitochondria and chloroplasts:
- Are surrounded by double phospholipid bilayers. The inner membrane of mitochondria is homologous to the plasma membrane of ancient aerobic bacteria, while the outer membrane is a relic of the host cell's engulfing vesicle.
- Contain their own circular DNA separate from the host cell nucleus, identical in structure to bacterial plasmids.
- Replicate via binary fission independently of the host cell cycle.
- Possess 70S ribosomes that are structurally similar to bacterial ribosomes (whereas the eukaryotic host's cytoplasm utilizes larger 80S ribosomes).
To make sense of this biological diversity, scientists use systematics, which is the scientific study of the diversity of organisms and their evolutionary relationships. The goal of systematics is to construct a phylogeny—a model that describes the evolutionary history and relationships of a species or group of species.
We visualize phylogenies using branching diagrams.
- A phylogenetic tree is a branching diagram representing evolutionary relationships among organisms. Crucially, the branch length of a phylogenetic tree can represent either the amount of evolutionary change or the chronological time elapsed.
- A cladogram is a specific type of phylogenetic tree showing branching order without proportional branch lengths. It shows who is related to whom, but not necessarily when the divergence occurred.
Anatomy of a Tree
To interpret these evolutionary models, you must fluently read their architecture:
- The root of a phylogenetic tree represents the most recent common ancestor of all entities in the tree.
- A node represents a speciation event separating a single ancestral lineage into descendant lineages.
- Terminal taxa are the specific organisms located at the very ends of the branches.
- Sister taxa are two lineages that are each other's closest evolutionary relatives. You can always spot them because sister taxa branch from the exact same node on a phylogenetic tree.
- A basal taxon is a lineage that diverges early in the history of a group and remains close to the root.
Clades and Groupings
Modern classification relies heavily on cladistics, a method of biological classification based entirely on evolutionary relationships and common ancestry. In cladistics, the gold standard is identifying a clade: a group of organisms that includes a single common ancestor and all of its descendants.
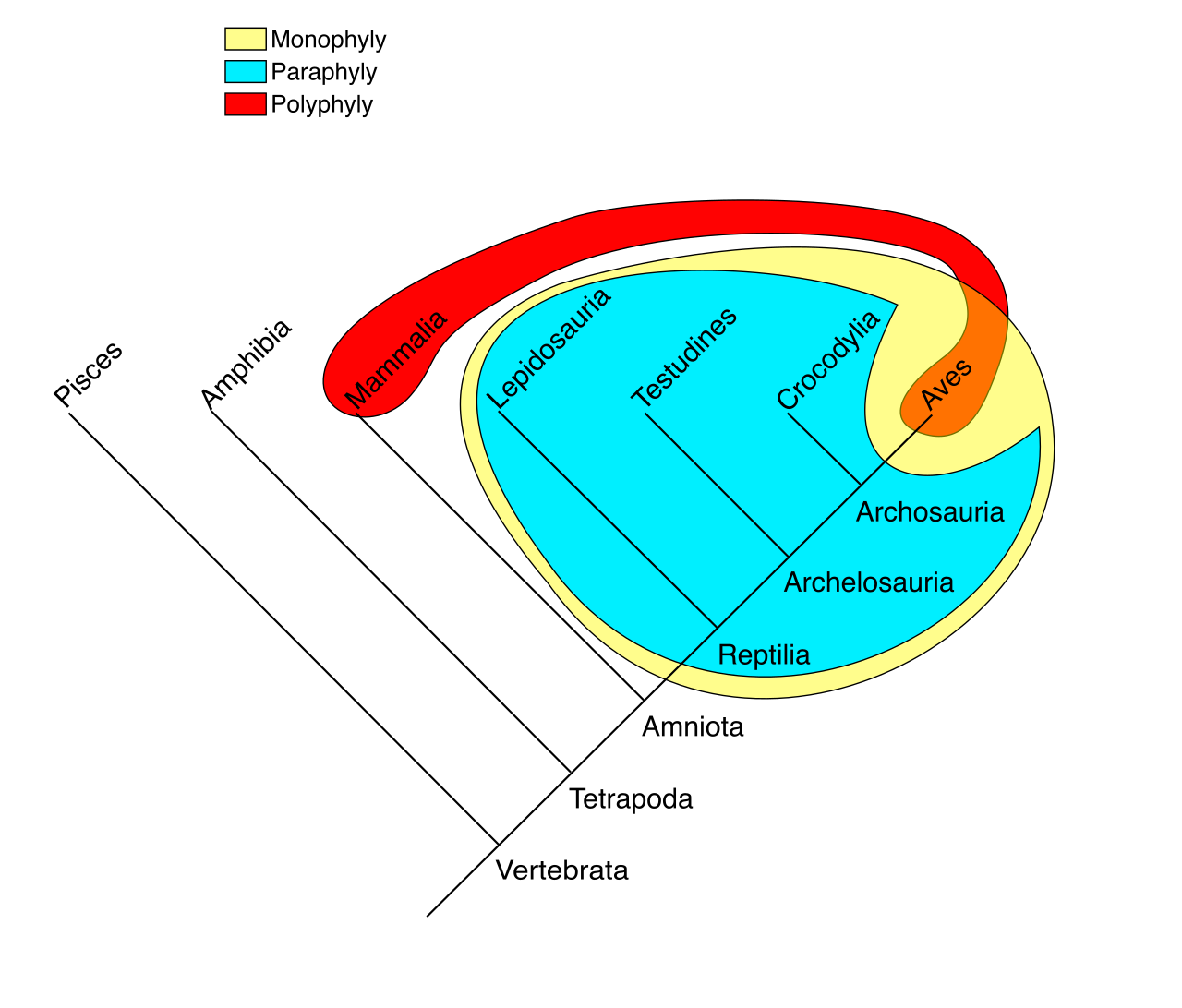
You will need to distinguish between three types of groupings:
| Grouping Type | Definition | Status in Cladistics |
|---|---|---|
| Monophyletic | A monophyletic group is perfectly equivalent to a clade. It includes an ancestor and all its descendants. | Valid |
| Paraphyletic | A paraphyletic group consists of a common ancestor and only a fraction of its descendants. (e.g., "Reptiles" traditionally excluded birds, making it paraphyletic). | Invalid |
| Polyphyletic | A polyphyletic group includes organisms with mixed evolutionary origins and entirely excludes the most recent common ancestor of its classified members. (e.g., grouping bats and birds together as "flying vertebrates"). | Invalid |
Constructing the Tree: Traits and Parsimony
How do scientists know where to draw the branches? By analyzing traits.
- A shared ancestral character is a trait that originated in an ancestor of the taxon (e.g., the backbone in mammals; fish also have backbones, so it predates mammals).
- A shared derived character is an evolutionary novelty unique to a particular clade (e.g., hair in mammals). Shared derived characters are also scientifically known as synapomorphies. Synapomorphies define clades.
To figure out whether a trait is ancestral or derived, systematists use an outgroup, which is a lineage known to have diverged before the lineage containing the specific groups being studied.
Because evolution can be messy (convergent evolution can cause similar traits to appear independently), trees are often constructed using the principle of maximum parsimony. Maximum parsimony assumes the most likely evolutionary pathway requires the fewest total evolutionary events. If Tree A requires feathers to evolve once, and Tree B requires feathers to evolve three separate times, Tree A is more parsimonious and highly favored.
While relative dating and cladograms give us the sequence of evolutionary events, modern biology uses DNA to estimate the exact timing of these divergences.
A molecular clock uses the mutation rate of biomolecules to deduce the prehistoric time when life forms diverged. How does this work? Molecular clocks rely on the assumption that certain non-coding regions of genomes mutate at a constant average rate. Because these regions do not code for critical proteins, mutations there are generally neutral and accumulate predictably over millions of years—like the steady ticking of a metronome.
If we know that a specific sequence mutates at a rate of one base pair per million years, and we see 10 differences between species A and species B, we can deduce they diverged roughly 10 million years ago. However, clocks run at different speeds in different genes. Therefore, molecular clocks are typically calibrated using known evolutionary divergence dates derived from the fossil record. By anchoring the molecular data to the absolute dates provided by radiometric dating of fossils, biologists bridge the gap between the rocks beneath our feet and the code within our cells.