Animal Evolution and Development
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Every animal on Earth, from the sponge filtering ocean currents to the high school student sitting in your biology classroom, begins as a single cell. The transformation of that solitary, unremarkable-looking sphere into a highly coordinated organism—equipped with a beating heart, a searching eye, and a thinking brain—is not a matter of vitalistic magic, but of exquisite mechanical and genetic choreography. For a biology educator, understanding this sequence is not merely about memorizing the developmental stages of a frog embryo; it is about grasping the underlying logic of the animal kingdom. By mastering the evolutionary trends and developmental cascades that shape an organism, you possess the conceptual scaffolding required to show your students how a simple flatworm and a complex mammal share the exact same evolutionary blueprints.
To understand animal complexity, we must first look at how life organizes itself. The biological hierarchy in animals progresses from cells to tissues to organs to organ systems to whole organisms. A single muscle cell contracts; a tissue of muscle cells generates force; a heart (an organ) pumps blood; the cardiovascular system distributes it; and the organism survives.
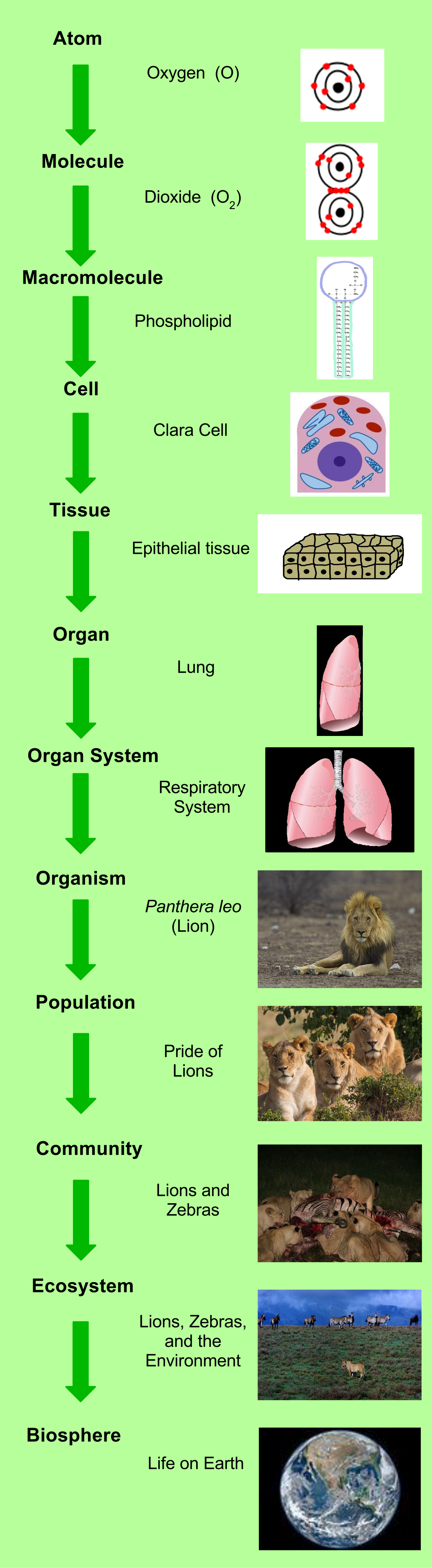
At the foundational level, animal tissues are classified into four primary categories:
- Epithelial tissue: Protects, secretes, and absorbs.
- Connective tissue: Supports and binds other tissues.
- Muscle tissue: Generates mechanical force.
- Nervous tissue: Transmits electrical signals.
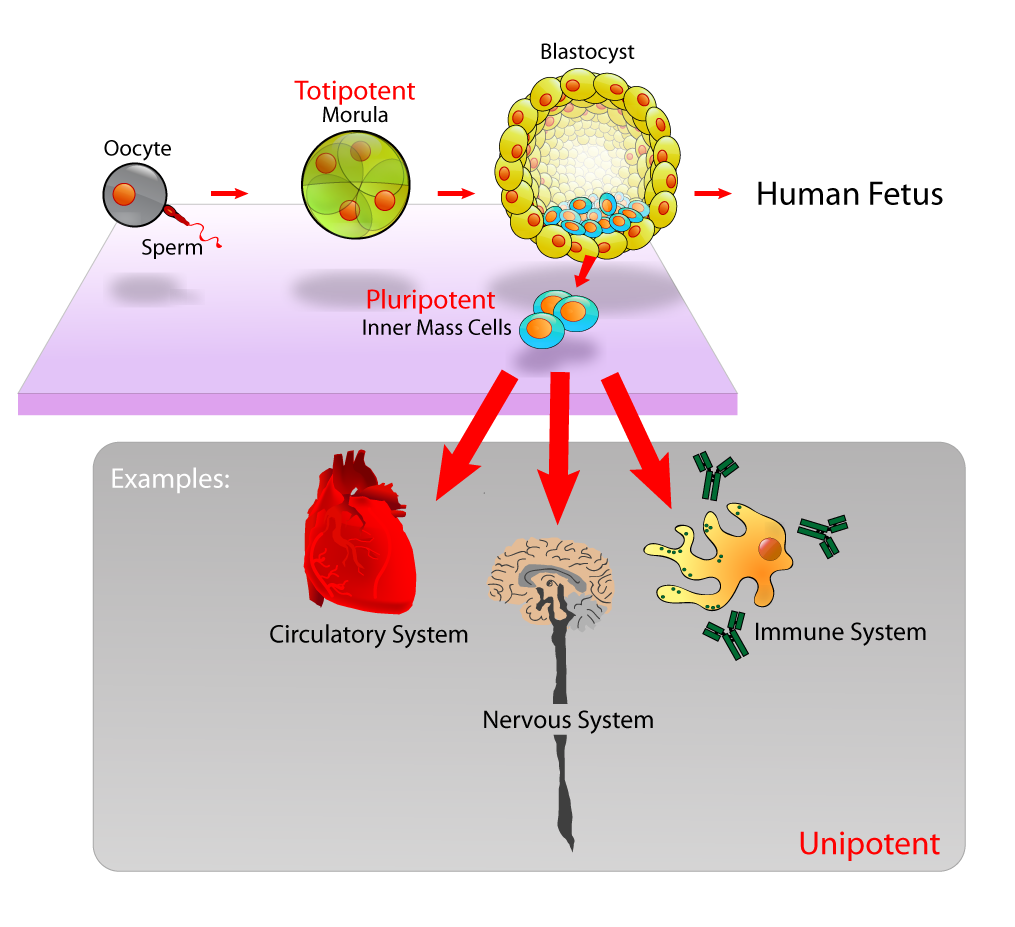
How Does a Cell Decide Its Fate?
A common misconception among students is that as cells specialize, they discard the DNA they no longer need. This is false. Cell differentiation is driven by the regulation of specific gene expression patterns rather than the loss of genetic material. Cell differentiation is the process by which unspecialized cells acquire specific structures and functions. Every cell has the entire "library" of the organism's DNA, but a liver cell only reads the liver chapters, while a neuron reads the brain chapters.
The raw material for this process comes from stem cells, which are undifferentiated cells capable of dividing and differentiating into specialized cell types. We categorize stem cells by their developmental potential:
- Totipotent stem cells are the ultimate blank slates. They can differentiate into any cell type within an organism and, crucially, are uniquely capable of giving rise to extraembryonic placental tissues.
- Pluripotent stem cells are slightly more restricted. They can differentiate into any cell type of the three embryonic germ layers, but cannot form the placenta.
- Multipotent stem cells are heavily specialized. They are restricted to forming specific cell lineages within a particular tissue type (e.g., hematopoietic stem cells in bone marrow can only become various types of blood cells).
The progression of animal evolution is a story of increasing structural organization. We categorize animal phyla based on profound structural differences that arise in embryonic development.
Symmetry and the Head
At the base of the animal tree sit the sponges in the phylum Porifera, which lack true differentiated tissues and generally exhibit body asymmetry. They are a loose confederation of cells without a definitive shape.
As we move up the evolutionary tree, distinct body plans emerge:
- Radial symmetry is a body plan where body parts are arranged radially around a central axis. Think of a bicycle wheel or a pie. This allows an animal to meet its environment equally from all sides, and radial symmetry is a primary characteristic of animals in the phylum Cnidaria (jellyfish, sea anemones).
- Bilateral symmetry is a body plan featuring distinct left and right mirror-image halves. With this evolutionary innovation came directed, forward movement. If an animal is always moving forward, it makes sense to put the sensory equipment and mouth at the front. Thus, the evolutionary emergence of cephalization is strongly associated with bilateral symmetry. Cephalization is the evolutionary concentration of sensory organs and nervous tissue at the anterior end of an animal.

Germ Layers
As animal embryos develop, they form distinct cellular layers that will eventually become tissues.
- Diploblastic animals develop from exactly two embryonic germ layers. The two germ layers in diploblastic animals are the ectoderm and the endoderm. (Cnidarians are diploblastic).
- Triploblastic animals develop from exactly three embryonic germ layers. The three germ layers in triploblastic animals are the ectoderm, mesoderm, and endoderm. All bilaterally symmetrical animals are triploblastic.
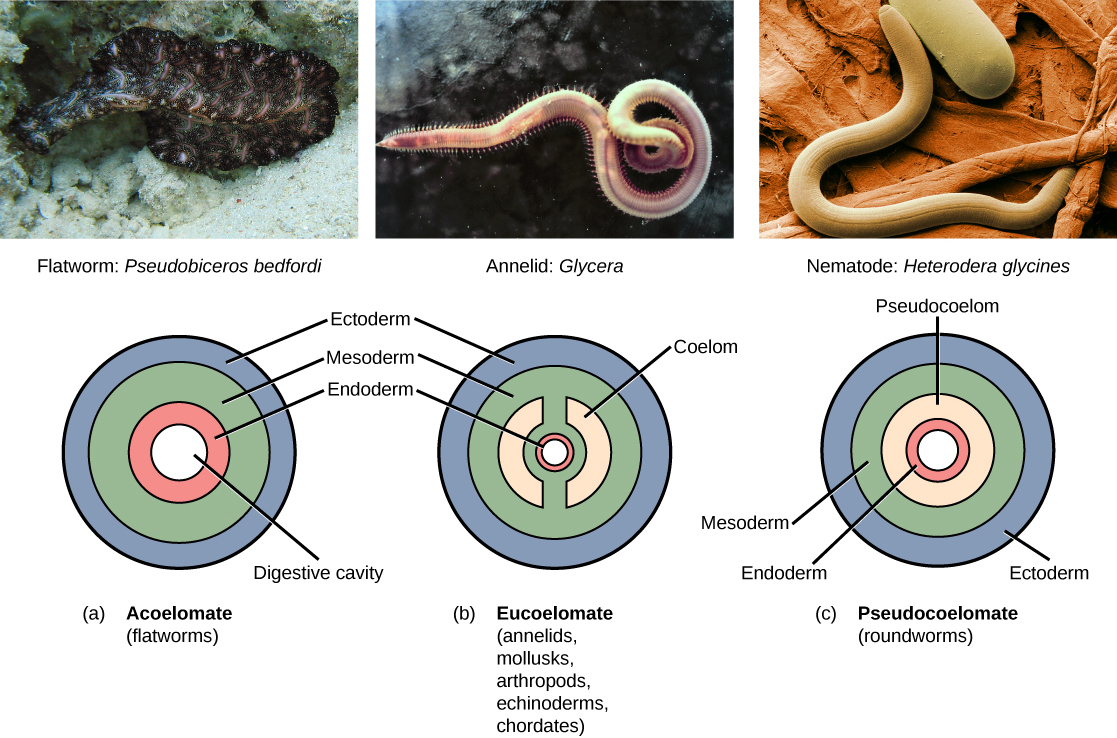
With the evolution of the mesoderm in triploblasts came the possibility of internal space.
Coelom: A fluid-filled body cavity located between the digestive tract and the outer body wall.
A coelom acts as a hydrostatic skeleton, cushions internal organs, and allows the digestive tract to move independently of the outer body wall. We classify triploblastic animals by their cavity architecture:
| Cavity Type | Definition | Example Phylum |
|---|---|---|
| Acoelomate | Animals that completely lack a fluid-filled body cavity between the digestive tract and the body wall. Everything is packed solid with tissue. | Flatworms in the phylum Platyhelminthes are classified as acoelomate animals. |
| Pseudocoelomate | Animals that possess a body cavity lined partially by mesoderm and partially by endoderm. | Roundworms in the phylum Nematoda are classified as pseudocoelomate animals. |
| Eucoelomate | Animals that possess a true coelom completely lined by mesoderm tissue. | Annelids, Mollusks, Arthropods, Echinoderms, Chordates. |
Protostomes and deuterostomes represent two major evolutionary lineages of bilaterally symmetrical animals. This divergence happens at the earliest stages of microscopic development and defines the structural fate of the organism.
The differences lie in cleavage (how the cells divide) and the fate of the blastopore (the first embryonic opening):
- Cleavage Patterns: Protostomes typically exhibit spiral and determinate cleavage during early embryonic development. This means the cell fate is locked in immediately; if you remove one cell, the embryo dies. Conversely, deuterostomes typically exhibit radial and indeterminate cleavage during early embryonic development. This indeterminate cleavage is what makes identical twins possible in humans—a separated early cell retains the capacity to form a whole organism.
- The Blastopore: The embryonic blastopore develops into the mouth in protostome animals (from the Greek proto = first, stoma = mouth). In contrast, the embryonic blastopore develops into the anus in deuterostome animals (deutero = second).
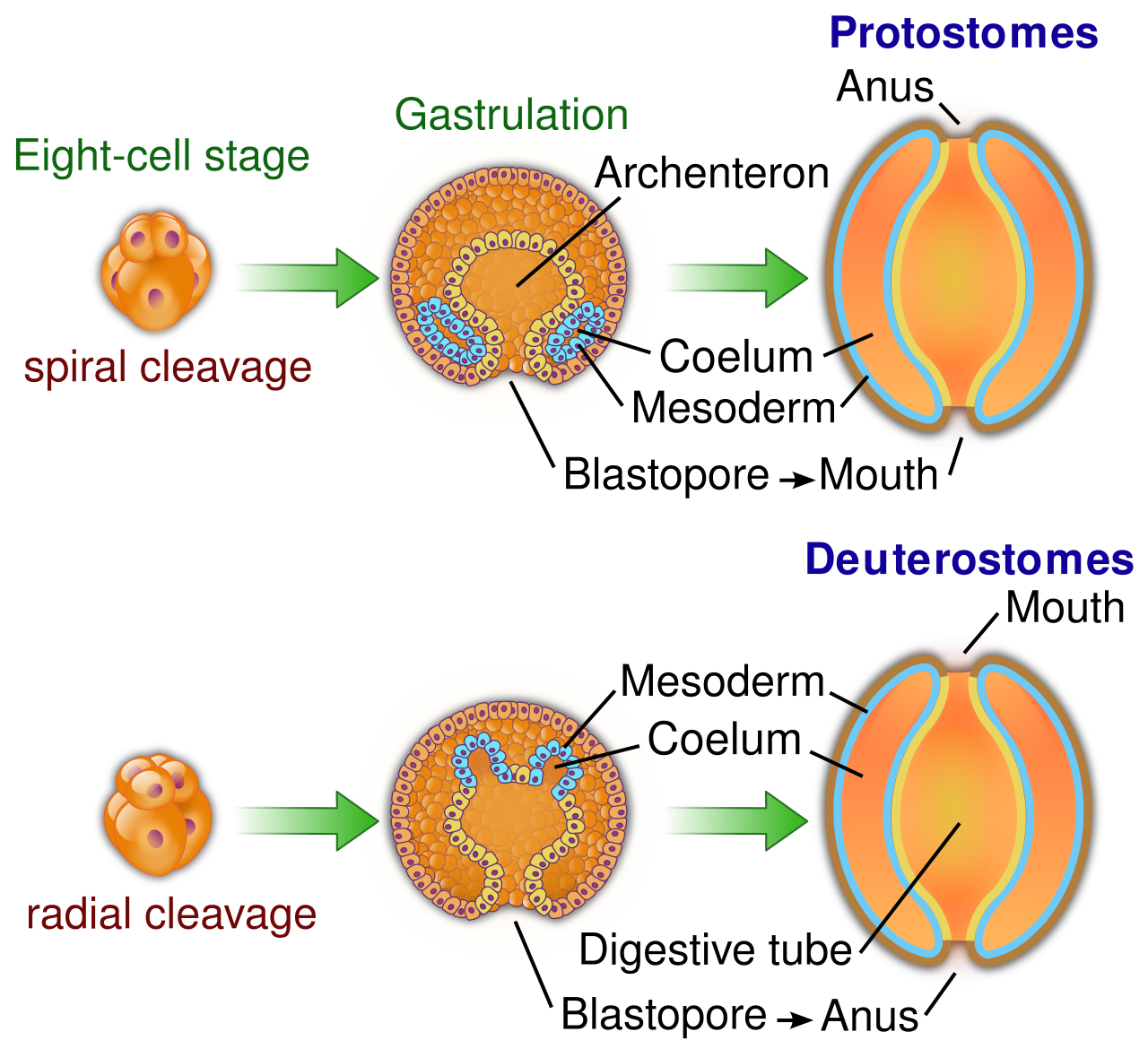
While a vast array of invertebrates are protostomes, animals in the phyla Echinodermata and Chordata are classified as deuterostomes. We humans are chordates; thus, structurally speaking, we share fundamental developmental mechanics with starfish.
To trace the development of an individual, we must start at conception. The vast majority of animal species are diploid organisms that reproduce sexually. To maintain the correct chromosome number across generations, animal gametogenesis utilizes the process of meiosis to produce haploid reproductive cells.
- Spermatogenesis is the specific biological process that produces male haploid gametes (sperm).
- Oogenesis is the specific biological process that produces female haploid gametes (eggs or ova).
Fertilization occurs when a haploid sperm cell successfully fuses with a haploid egg cell. The biological process of fertilization produces a single diploid cell called a zygote.
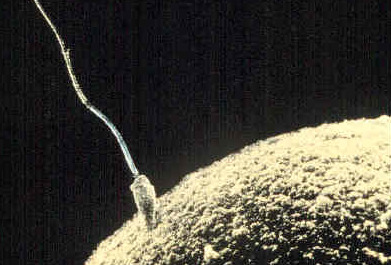
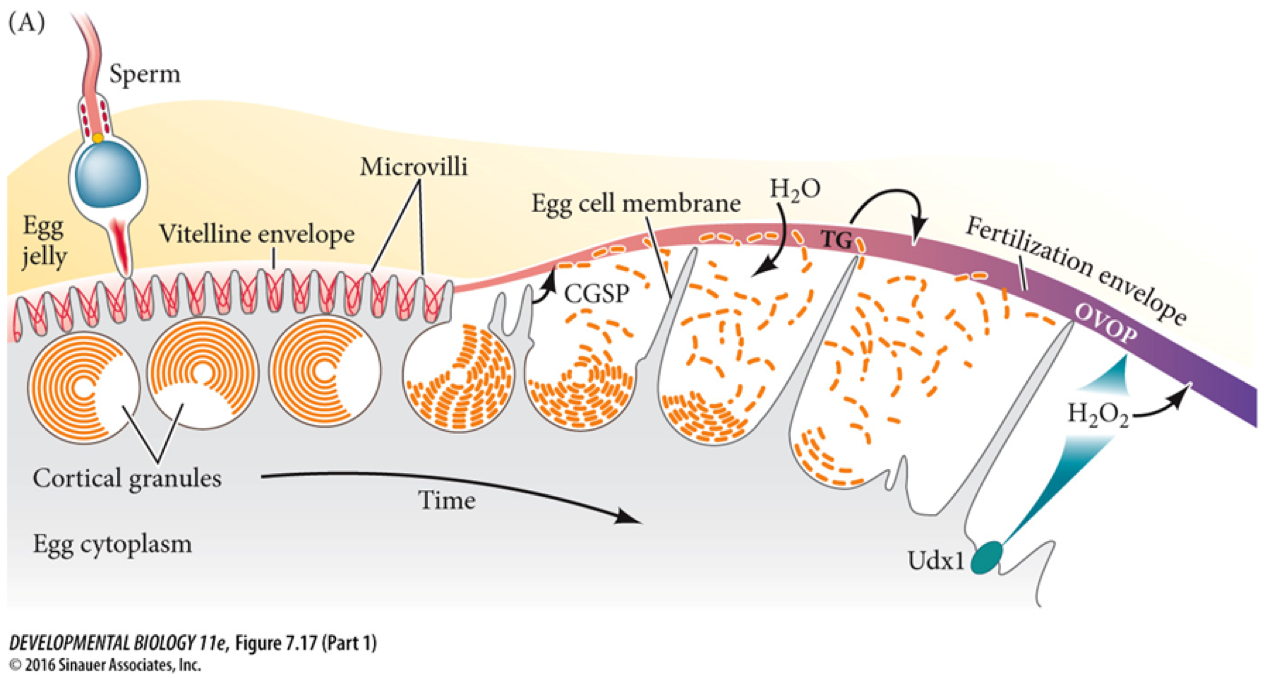
Blocking Polyspermy
If multiple sperm fuse with one egg (polyspermy), the zygote receives too many chromosomes and inevitably dies. The egg defends itself using two sequential mechanisms:
- The Fast Block: The moment the first sperm contacts the egg, sodium ion channels open. The fast block to polyspermy involves the rapid electrical depolarization of the egg cell membrane. This electrical shock lasts only a few minutes, acting as a temporary shield.
- The Slow Block: The electrical depolarization triggers a massive release of calcium ions inside the egg, causing the slow block to polyspermy, which is initiated by a mechanism called the cortical reaction. The cortical reaction physically alters the egg's extracellular matrix to prevent additional sperm from entering, turning the soft outer layer into an impenetrable fertilization envelope.
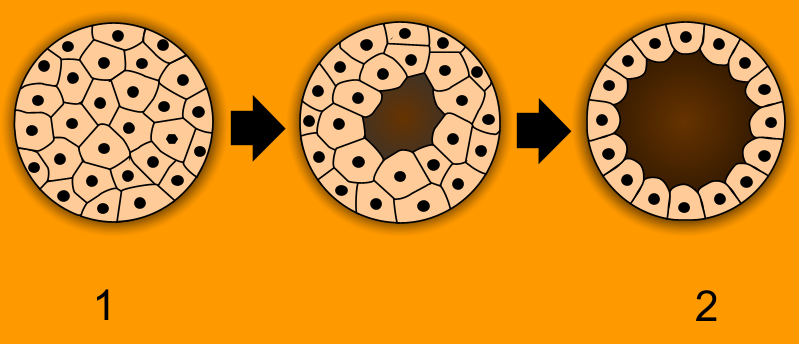
Once the zygote is formed, it must quickly build a multicellular body.
Cleavage is a series of rapid mitotic cell divisions that immediately follow fertilization. A profound characteristic of this stage is that embryonic cleavage divisions occur without any overall increase in the total volume of the embryo. The massive zygote simply chops itself into smaller and smaller cells, skipping the typical growth phases of the cell cycle.
- After several rounds of cleavage, the embryo becomes a morula, which is a solid ball of embryonic cells.
- As divisions continue, cells pump fluid into the center, forming a hollow sphere. A blastula is a hollow ball of embryonic cells formed from the continuous division of the morula.
- The blastocoel is the central fluid-filled cavity located inside the blastula. (Note for educators teaching human biology: In mammalian development, the embryonic blastula stage is specifically referred to as a blastocyst, which contains the inner cell mass that will eventually become the fetus).
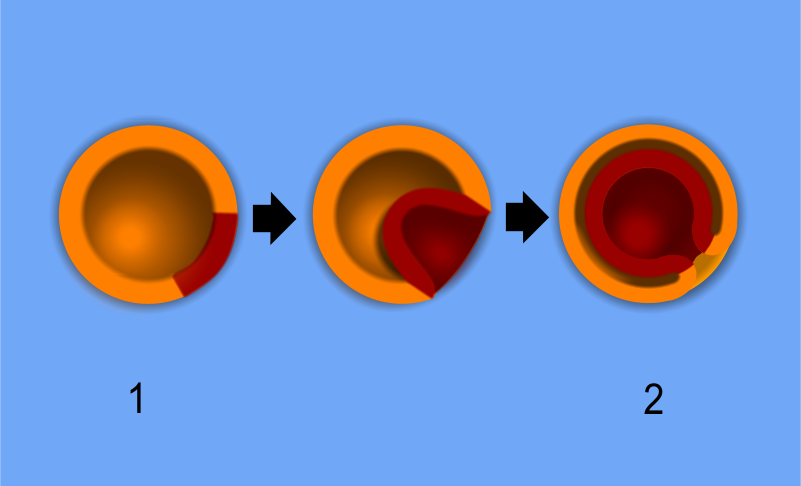
Gastrulation: The Most Important Time in Your Life
The biologist Lewis Wolpert famously said, "It is not birth, marriage, or death, but gastrulation which is truly the most important time in your life."
Gastrulation is the developmental process where blastula cells actively migrate and rearrange to form embryonic germ layers. Imagine taking a hollow rubber ball (the blastula) and poking your thumb into it. That mechanical inward folding changes the 3D geometry of the entire organism.
- Gastrulation creates an inward cellular fold known as the archenteron.
- The blastopore is the distinct opening of the archenteron to the external environment. (Recall our discussion of protostomes and deuterostomes—this is where that opening forms).
- As development progresses, the archenteron eventually develops into the primitive digestive tract of the animal.
Following gastrulation, the embryo possesses its three germ layers. Next comes organogenesis, the developmental phase where the established embryonic germ layers differentiate into specific internal organs.
The mapping of these layers is highly conserved across triploblastic animals:
- The embryonic ectoderm germ layer develops into the nervous system and epidermal skin structures. (It makes sense: the outside layer becomes the outer skin and the sensors to interact with the outside world).
- The embryonic mesoderm germ layer develops into the muscular, skeletal, and circulatory systems. (The middle layer forms the structural bulk and transport highways).
- The embryonic endoderm germ layer develops into the internal linings of the digestive and respiratory tracts. (The inner layer coats the inner tubes).
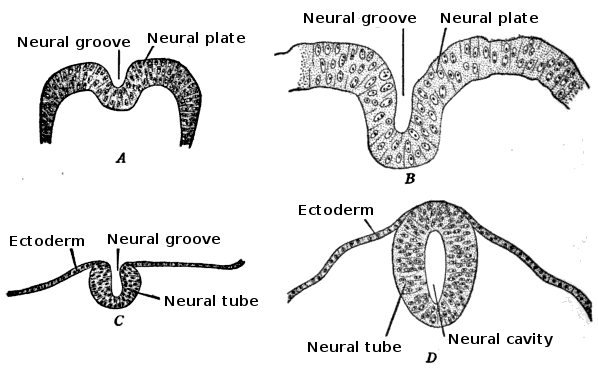
Neurulation
In chordates, a highly specific phase of early organogenesis occurs. Neurulation is the specific developmental process in chordate embryos that forms the neural tube.
How does the ectoderm "know" it is time to fold into a tube? It is chemically commanded by the mesoderm beneath it. The notochord is a specialized mesodermal rod that chemically induces the overlying ectoderm to form the neural plate. This plate then folds inward and pinches off. Ultimately, the embryonic neural tube eventually differentiates into the adult central nervous system (the brain and spinal cord).
Hox Genes: The Architects of the Body Plan
The precision of organogenesis is governed by master control switches known as homeotic genes. Specifically, Hox genes are a family of regulatory genes that control the body plan of an embryo.
These genes do not build the eye or the leg directly; rather, they serve as high-level executives. Hox genes encode specialized transcription factors that direct the formation of distinct anatomical body parts. They tell the developing cells "where" they are in 3D space. Thus, Hox genes play a critical role in establishing the anterior-posterior axis during animal development. If a Hox gene mutates, a fruit fly might grow legs out of its head instead of antennae—the structural genes work perfectly, but the master architect handed out the wrong blueprints for that specific location.
By bridging the molecular scale of transcription factors to the macroscopic scale of an animal's body plan, we come full circle. The beauty of teaching biology lies in revealing this continuity: the same genetic switches and folding tissue layers govern the development of virtually every animal your students will ever encounter.