Taxonomy and Microorganisms
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Imagine inheriting an uncatalogued library containing millions of books, spanning billions of years of authorship, written in a language you are just learning to read. That is the predicament of the biological sciences. To make sense of the dizzying diversity of life on Earth, we cannot merely list what exists; we must organize it into a framework that reflects the very history of its creation. For an aspiring biology educator, mastering this framework is not about memorizing a static dictionary of Latin terms. It is about equipping your future students with a map of evolutionary history—a way to see that every microbe, mushroom, and mammal is a surviving thread in an unbroken lineage stretching back to the dawn of life.
To navigate the library of life, biologists employ three distinct but deeply interwoven disciplines:
- Taxonomy is the scientific discipline of describing, naming, and classifying organisms.
- Phylogeny describes the evolutionary history and evolutionary relationships among a species or a group of species.
- Systematics is the biological field that organizes and classifies organisms based on evolutionary relationships.
Think of taxonomy as the labeling system, phylogeny as the family tree, and systematics as the grand endeavor that ensures our labels accurately reflect that tree.
Linnaeus and the Rules of Naming
Before the 18th century, species were named using long, chaotic descriptive phrases. Carl Linnaeus revolutionized this by developing the binomial nomenclature system for formally naming organisms.
Binomial nomenclature assigns each distinct organism a unique two-part scientific name.
In binomial nomenclature, the first word represents the genus name, and the second word represents the specific epithet or species name. To maintain universal clarity—a critical science and engineering practice—standard biological naming conventions require the genus name to be capitalized, while the species name must be written entirely in lowercase letters. Furthermore, scientific names formatted under binomial nomenclature must be italicized in print or underlined when handwritten (e.g., Homo sapiens or <u>Escherichia coli</u>).
The Taxonomic Hierarchy
The modern taxonomic classification system uses a hierarchical model. As you move down the hierarchy, the groupings become more exclusive. The eight primary taxonomic levels from most inclusive to least inclusive are domain, kingdom, phylum, class, order, family, genus, and species.
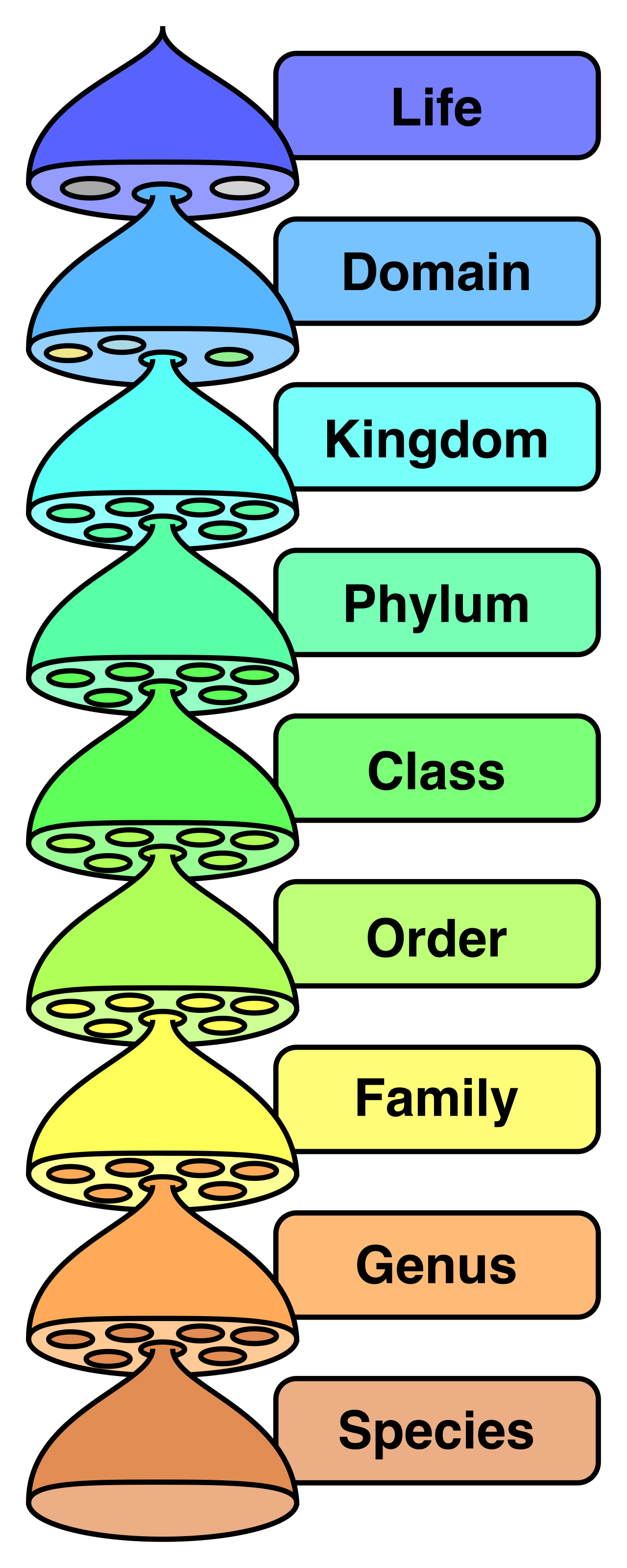
A taxon is a specific group of organisms at any given level within the taxonomic classification system. For example, Mammalia is a taxon at the class level.
For decades, students were taught the traditional six-kingdom biological classification system, which includes Eubacteria, Archaebacteria, Protista, Fungi, Plantae, and Animalia. However, as our molecular sequencing improved, we realized the fundamental split in life occurred much earlier and deeper. Today, the overarching classification rests on the three domains of life: Bacteria, Archaea, and Eukarya.
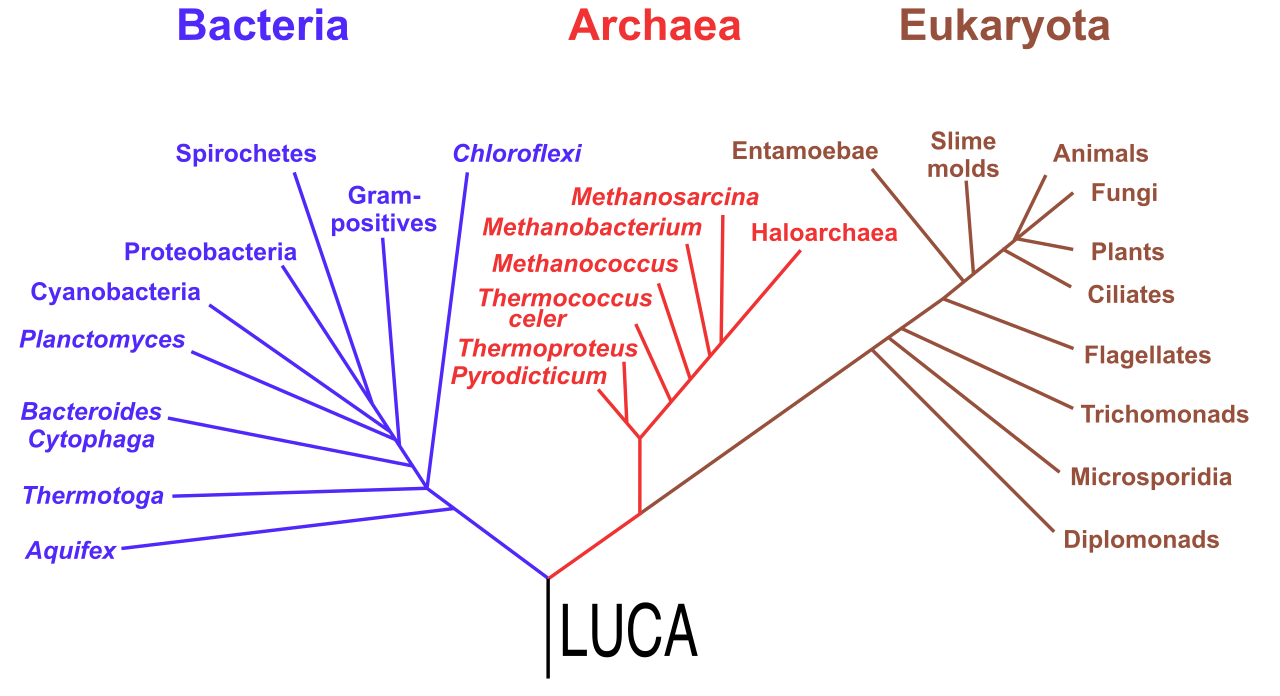
To determine how to group these domains and kingdoms, systematists use cladistics.
Cladistics classifies organisms into monophyletic groups based on shared derived characteristics.
When evaluating evolutionary trees with your students, you will frequently encounter three types of groupings:
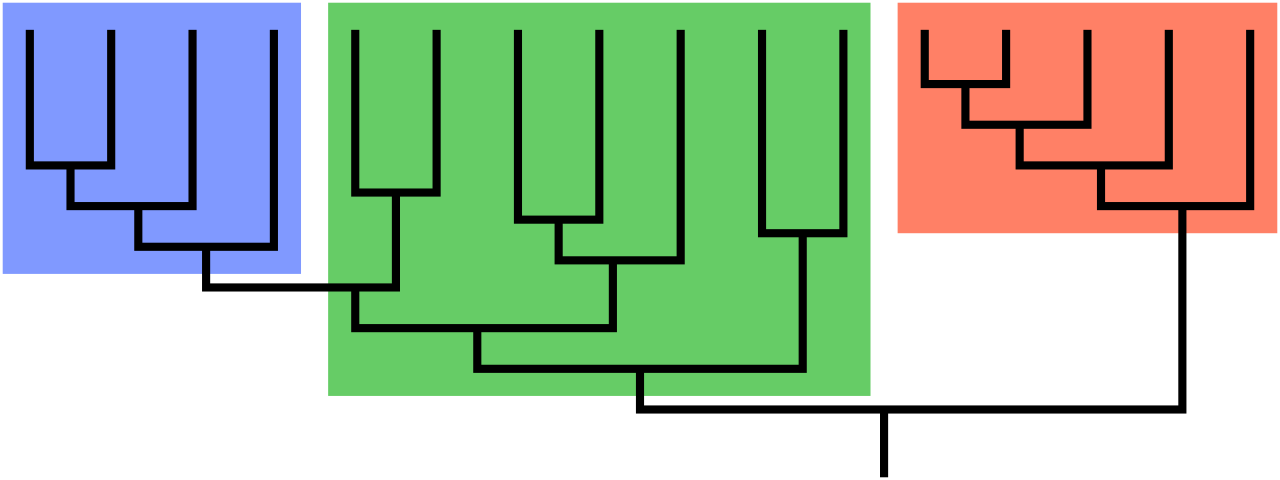
- A monophyletic clade consists of a single evolutionary ancestor and all the descendants of that ancestor. This is the gold standard of modern classification.
- A paraphyletic taxonomic group includes a common ancestor and only a subset of the descendants of that ancestor. (As we will see, reptiles and protists are classic examples).
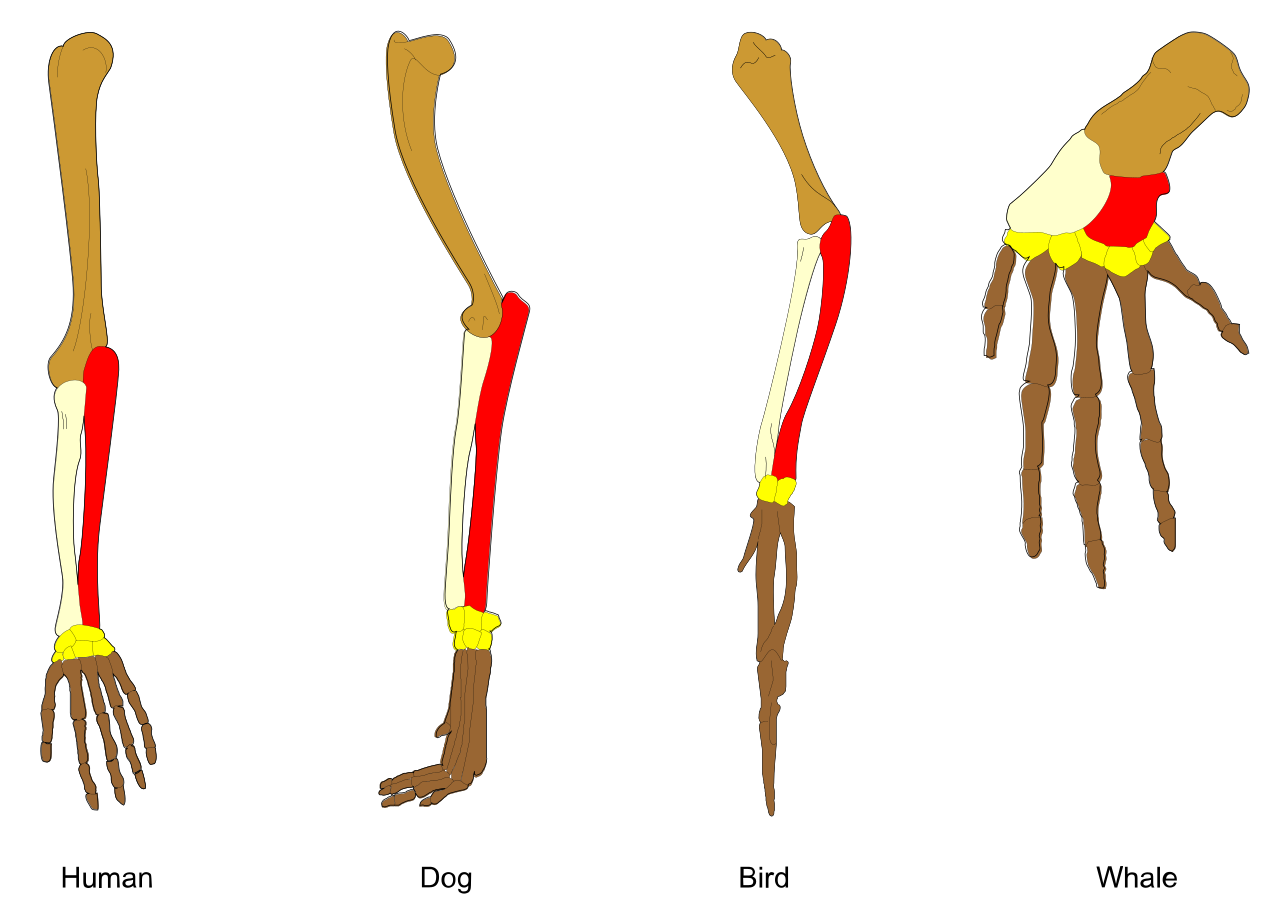
To build accurate trees, we must distinguish between features that look alike and features that actually share a genetic history. Homologous structures share a similar embryonic origin resulting from descent from a common evolutionary ancestor (like a human arm and a bat wing). In contrast, analogous structures perform similar functions due to convergent evolution rather than common ancestry (like a bat wing and an insect wing).
Before diving into the microorganisms, we must establish the baseline of cellularity.
- Unicellular microorganisms contain a single cell that executes all physiological functions required for survival.
- Multicellular microorganisms rely on differentiated, specialized cells cooperating to maintain the life of the entire organism.
All known species of Bacteria and Archaea exist exclusively as unicellular organisms. Eukarya, however, spans both unicellular and multicellular forms.
Viruses blur the line between the living and the inert. Viruses are acellular entities, meaning they are not composed of cells. Because they lack an independent cellular metabolism, they are obligate intracellular parasites requiring a host cell's machinery to synthesize proteins and replicate.
Viral Anatomy
A viral particle is stripped down to the bare minimum. It consists of a nucleic acid core enclosed within a protein coat called a capsid. The genetic material of a virus consists of either DNA or RNA, and unlike cellular life, viral genomes can be single-stranded or double-stranded.
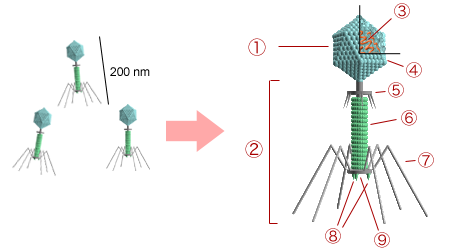
The capsid itself is built from individual protein subunits that assemble to form it, known as capsomeres. Furthermore, enveloped viruses possess an outer lipid envelope derived from the plasma membrane of a host cell—a molecular camouflage. Protruding from this structure are viral surface glycoproteins, which act like lockpicks; they bind to specific receptor molecules on the exterior of a host cell to gain entry.
Viral Replication Cycles
Bacteriophages are specialized viruses that infect bacterial cells. When a virus infects a host, it generally follows one of two paths:
- The lytic viral replication cycle results in the immediate rupture and death of the infected host cell as new viral particles burst forth.
- The lysogenic viral replication cycle is stealthier; it involves the integration of the viral genome into the host cell's chromosome, sitting dormant as the host divides.
Certain RNA viruses break the standard flow of genetic information (DNA → RNA → Protein). Retroviruses are RNA viruses that utilize the enzyme reverse transcriptase to synthesize a DNA copy from a viral RNA template. (HIV is the most prominent example to share with your students).
Subviral Agents
The biological world also contains pathogens even simpler than viruses:
- Prions are infectious pathogen particles composed entirely of misfolded proteins. They contain no detectable nucleic acids, yet they cause cascading neurological diseases (like Mad Cow Disease).
- Viroids are small infectious plant pathogens consisting solely of circular, single-stranded RNA without a protein coat.
Bacteria are unicellular prokaryotic organisms. Their defining anatomical feature is that bacterial cell walls are primarily composed of a polymer called peptidoglycan.
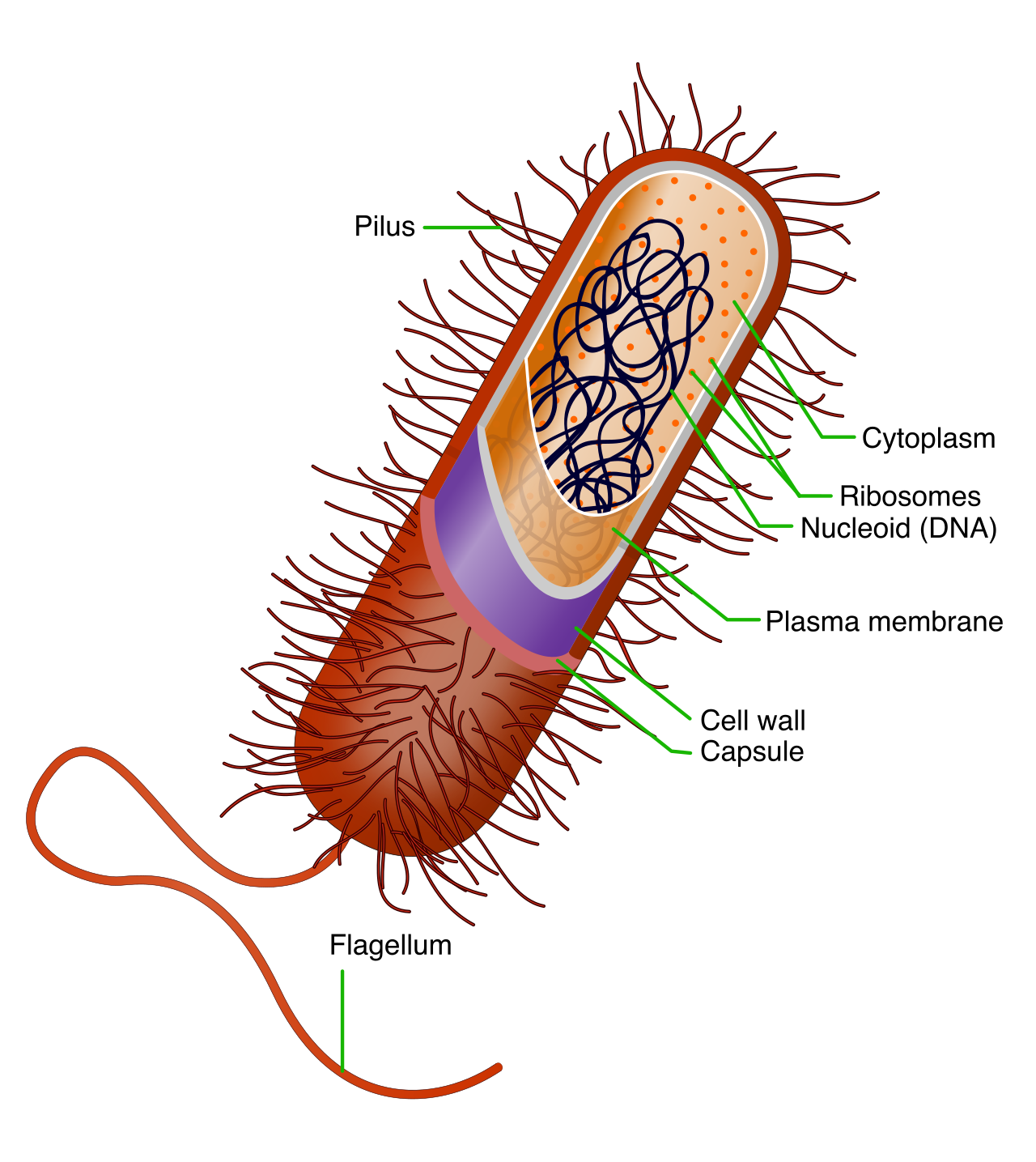
Inside the cell, the primary genetic material of a bacterium is typically a single circular chromosome. Because they lack a true nucleus, the bacterial chromosome is located in an irregularly shaped cytoplasmic region called the nucleoid.
Bacteria often carry auxiliary genetic backpacks called plasmids—small, circular DNA molecules existing independently of the main chromosomal DNA. Plasmids frequently carry accessory genes conferring advantageous traits such as antibiotic resistance.
Bacterial Morphology and Staining
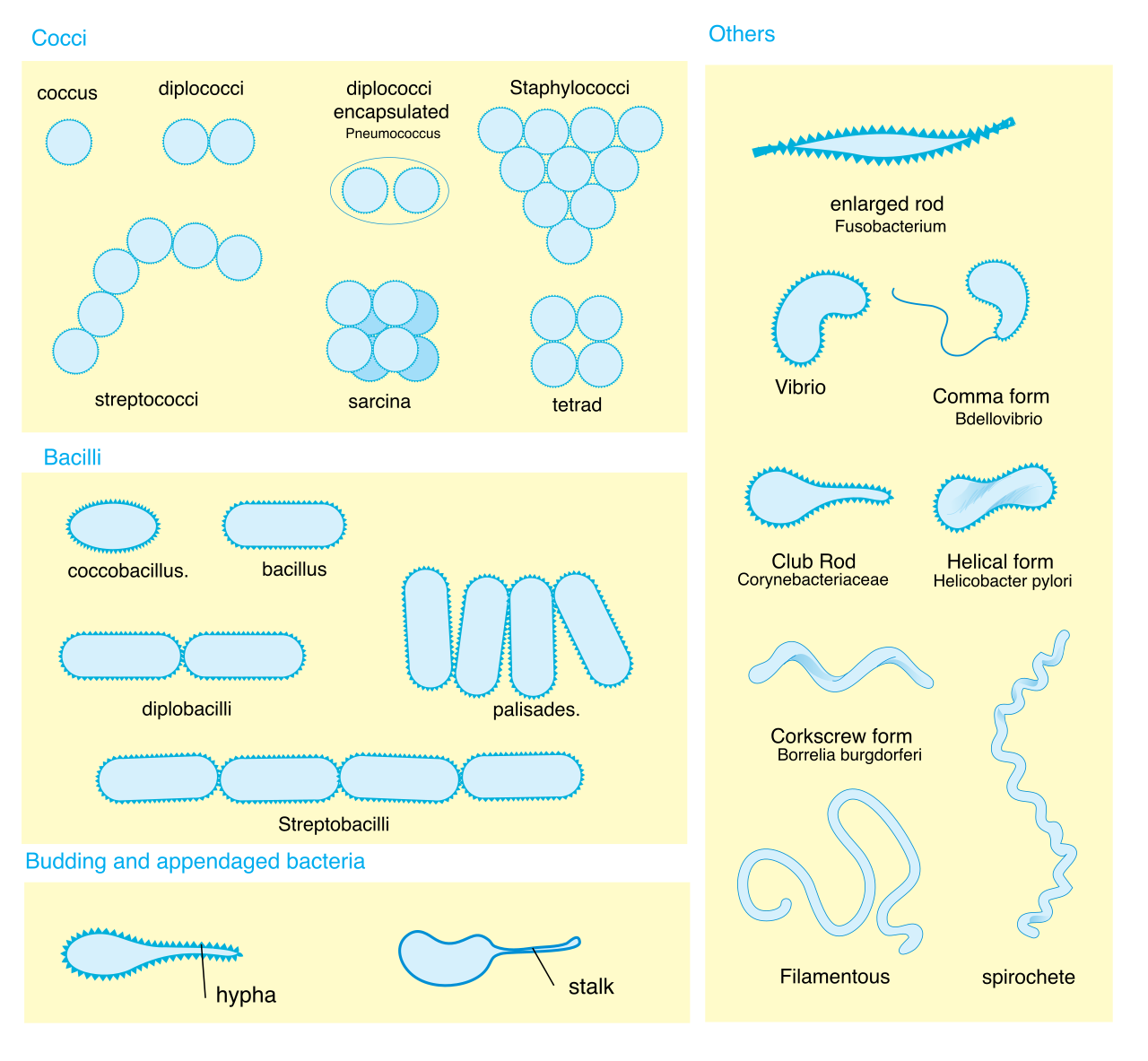
Under a microscope, bacteria generally exhibit three classic shapes:
- Cocci are spherically shaped bacterial cells.
- Bacilli are rod-shaped bacterial cells.
- Spirilla are spiral-shaped bacterial cells.
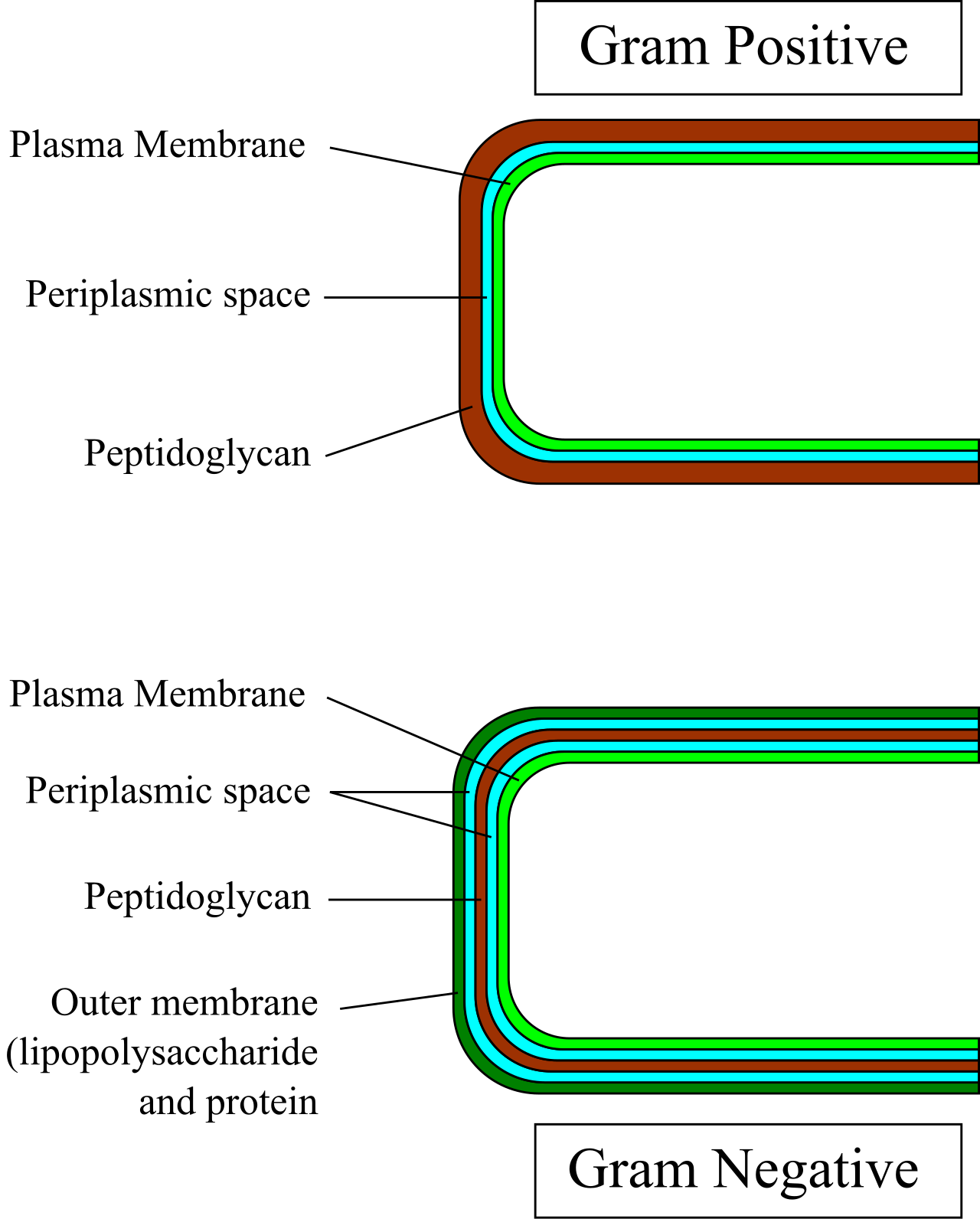
In the lab, scientists classify bacteria using a Gram stain, which detects cell wall architecture:
- Gram-positive bacteria possess a remarkably thick cell wall composed of many overlapping layers of peptidoglycan. They retain crystal violet dye and appear purple after a standard Gram stain.
- Gram-negative bacteria possess a thin layer of peptidoglycan located between the inner plasma membrane and an outer lipid membrane. Because of this thin layer, they lose the crystal violet dye and appear pink or red after counterstaining.
Bacterial Physiology and Reproduction
The predominant mode of asexual reproduction shared by both Bacteria and Archaea is binary fission. This process results in the division of a single parent cell into two genetically identical daughter cells.
Yet, bacteria evolve rapidly through horizontal gene transfer, which occurs in three ways:
- Bacterial transformation occurs when a live bacterium absorbs fragments of naked DNA from the surrounding extracellular environment.
- Bacterial transduction occurs when a bacteriophage accidentally transfers genetic material from a previously infected bacterium to a newly infected bacterium.
- Bacterial conjugation involves the direct physical transfer of genetic material from a donor bacterium to a recipient bacterium via a specialized structure called a pilus.
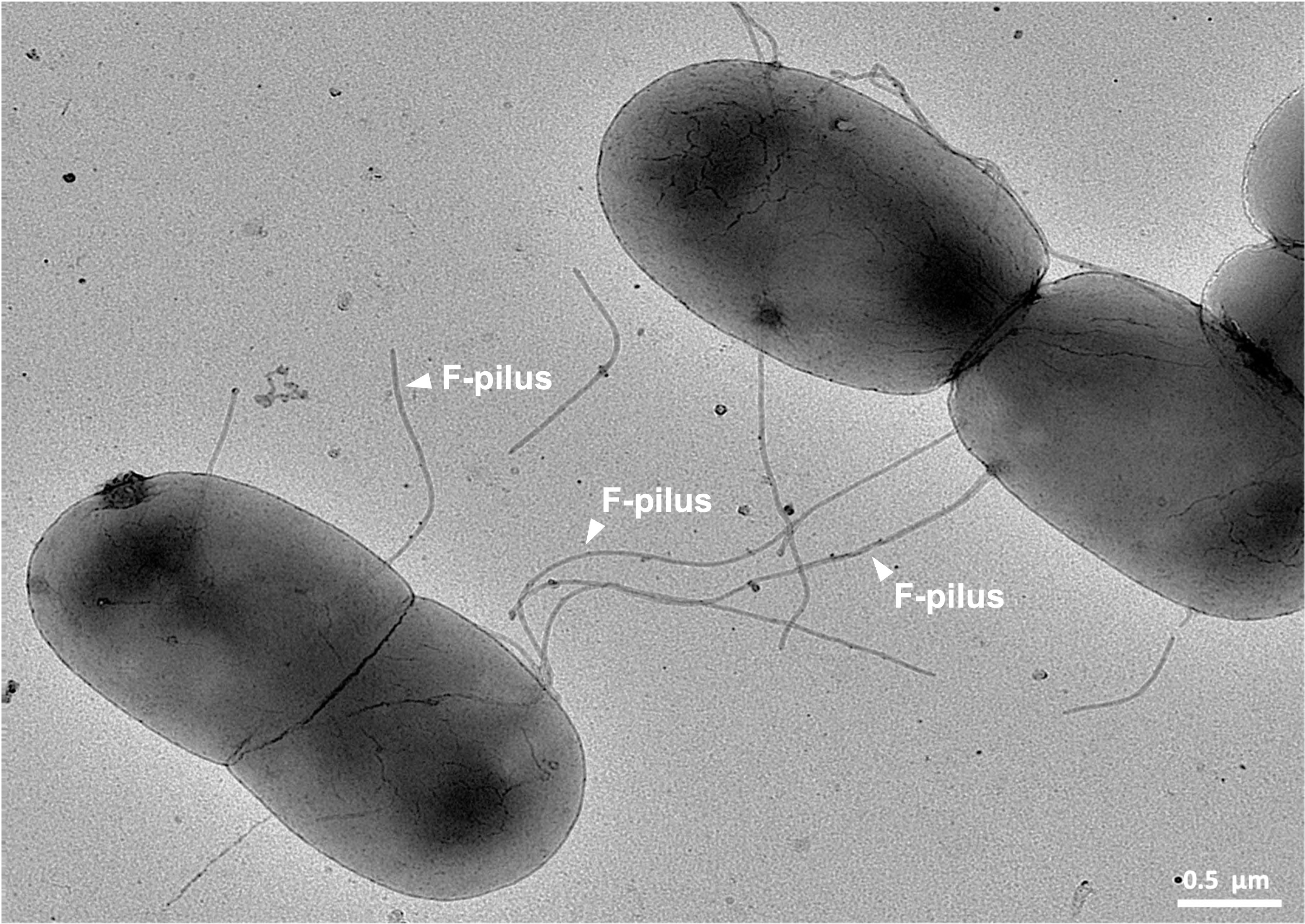
Some bacteria are crucial to our planet's energy flows. Cyanobacteria are photoautotrophic bacteria. Billions of years ago, they transformed our atmosphere because they generate oxygen gas as a direct metabolic byproduct of photosynthesis.
When conditions turn lethal, some bacteria deploy the ultimate survival mechanism: endospores. Endospores are highly durable, dormant bacterial structures capable of surviving extreme environmental stressors like boiling, radiation, and desiccation.
Originally mistaken for bacteria, Archaea are unicellular prokaryotic organisms with profound biochemical differences. Most notably, the cell walls of Archaea completely lack peptidoglycan. Instead, many archaeal species possess cell walls constructed from a polymer known as pseudopeptidoglycan.
Their cell membranes are equally unique. The plasma membranes of Archaea contain ether linkages between the glycerol molecule and the lipid chains, and archaeal lipid chains frequently exhibit branched molecular structures. In contrast, the plasma membranes of Bacteria and Eukarya contain ester linkages with unbranched chains.
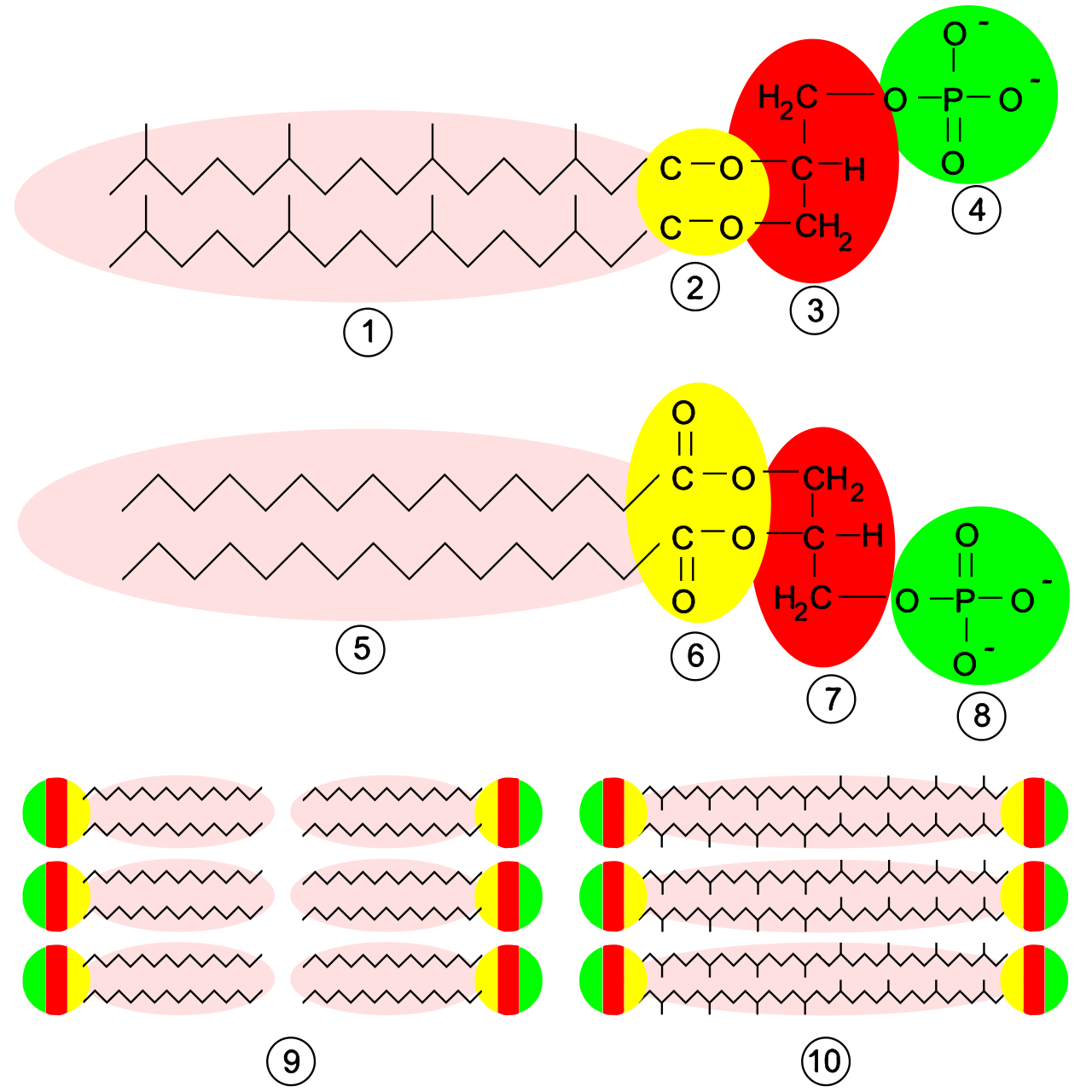
Fascinatingly, despite being prokaryotes, the transcription and translation molecular machinery in Archaea more closely resembles the machinery in Eukarya than the machinery in Bacteria.
The Edge of Life
Many Archaea are extremophiles—organisms that achieve optimal growth in physically or geochemically extreme environmental conditions.
- Halophiles are extreme archaea that require highly saline environments for survival (like the Dead Sea).
- Thermophiles thrive at exceptionally high temperatures (like hydrothermal vents).
- Methanogens are archaea that generate methane gas as a byproduct of cellular metabolism. They are obligate anaerobes, meaning they are organisms that undergo lethal cellular damage in the presence of oxygen.
Protists are the ultimate biological grab-bag. Protists are organisms classified within the domain Eukarya, meaning protist cells contain a distinct membrane-bound nucleus protecting the cellular DNA.
Because they represent such a diverse array of lineages, the kingdom Protista is widely recognized by modern taxonomists as a paraphyletic group—they do not form a single monophyletic clade.
While the majority of protist species are unicellular organisms, there are spectacular exceptions; giant kelps are an example of large multicellular protists forming vast underwater forests.
Protist Nutrition and Motility
Protists are incredibly versatile in how they secure energy:
- Photoautotrophic protists synthesize organic compounds utilizing chloroplasts.
- Heterotrophic protists obtain organic carbon exclusively by consuming other organisms or organic matter.
- Mixotrophic protists can alternate between photoautotrophic and heterotrophic nutritional modes based on current environmental conditions.
Heterotrophs often feed via phagocytosis, an active transport process involving the cell membrane engulfing a solid particle to form an intracellular food vacuole.
To find food, protists utilize diverse motility mechanisms:
- Cilia: Certain protist species exhibit motility propelled by short, hair-like cellular appendages.
- Flagella: Other species exhibit motility propelled by long, whip-like cellular structures.
- Pseudopodia: Amoeboid protists move by temporarily extending portions of cytoplasm called pseudopodia ("false feet").
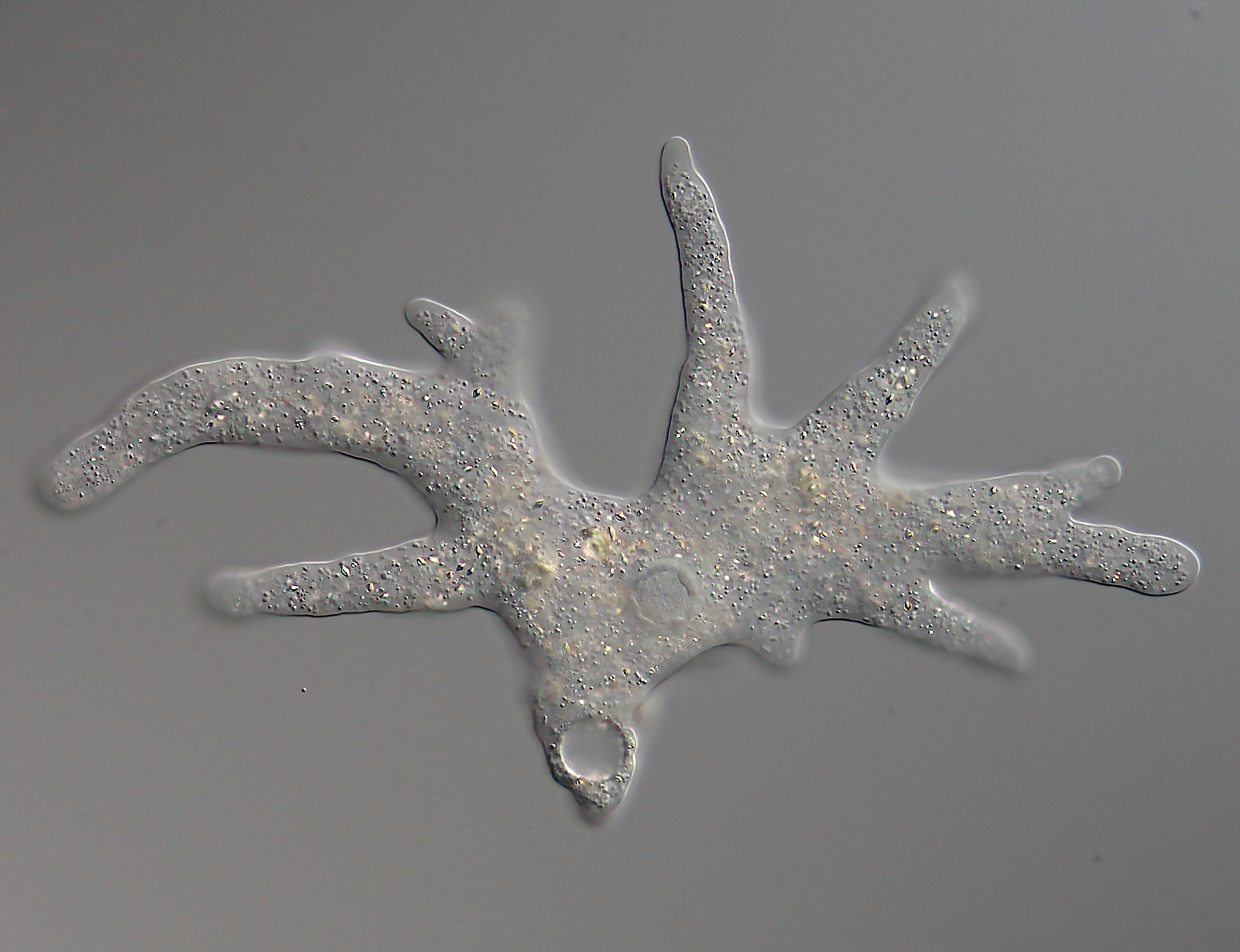
Diversity and Reproduction
Notable protist groups include:
- Diatoms: Unicellular photosynthetic protists encased in protective distinct cell walls composed of silica (glass).
- Dinoflagellates: Marine and freshwater protists structurally characterized by the presence of two flagella.
While many reproduce asexually, specialized sexual reproduction exists. For instance, sexual reproduction in paramecia occurs through conjugation involving the direct exchange of haploid micronuclei between two temporarily joined protist cells. (Do not confuse this with bacterial conjugation, which involves transferring a plasmid via a pilus!).
Fungi might look like plants, but they are genetically closer to animals. Fungi are non-photosynthetic organisms classified within the domain Eukarya.
All fungi are obligate heterotrophic organisms. Unlike humans who digest food internally, fungi execute external digestion. They acquire nutrients by secreting digestive exoenzymes into their external environment and subsequently absorbing the broken-down organic molecules. Many are vital ecosystem recyclers; saprobic fungi derive essential nutrients from decaying, dead organic matter.
Fungal Anatomy
Unlike plant cell walls made of cellulose, fungal cell walls are structurally reinforced by a complex polysaccharide named chitin (the same material in a beetle's exoskeleton).
The majority of identified fungal species are multicellular organisms. The vegetative body of a multicellular fungus consists of long, thread-like filament structures called hyphae. When these hyphae weave together into a dense, interconnected network, the structure is formally termed a mycelium.
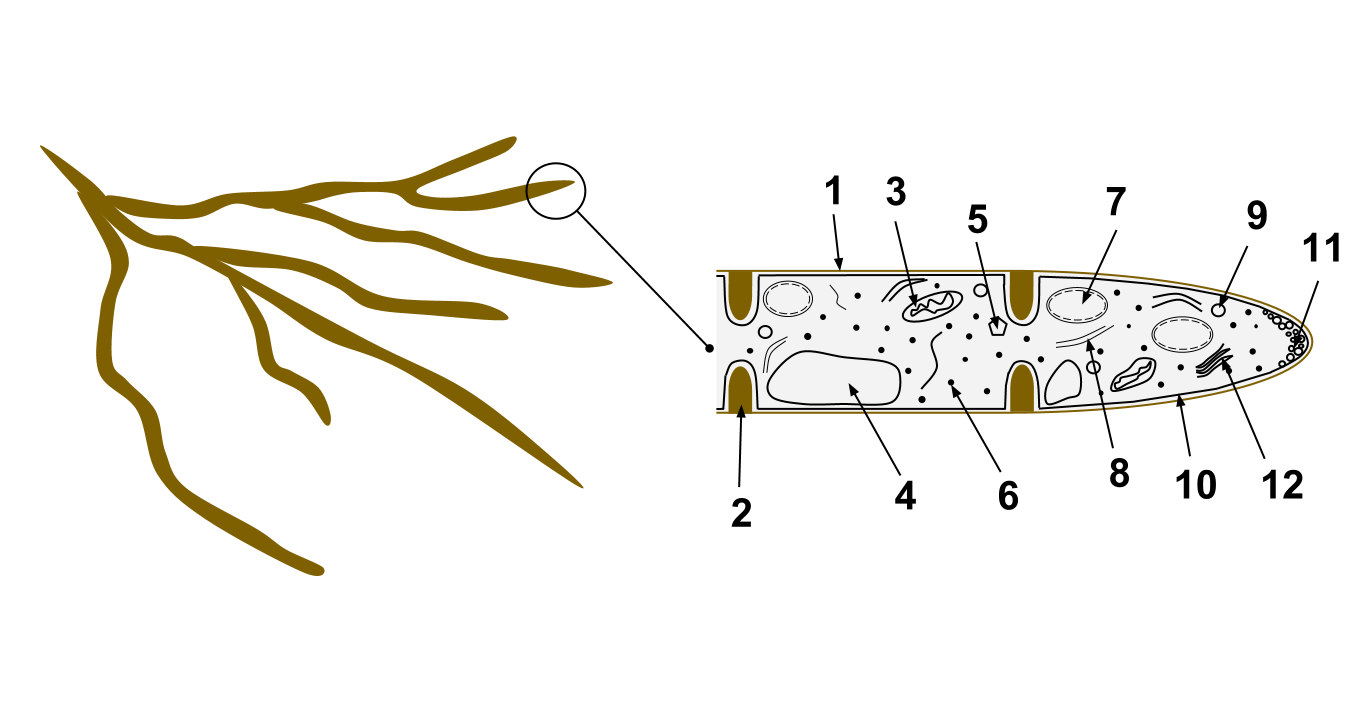
Hyphae come in two structural variants:
- Septate hyphae are physically divided into separate cellular compartments by internal cross-walls called septa.
- Coenocytic hyphae completely lack internal septa. As a result, they form a single, continuous cytoplasmic mass containing hundreds or thousands of nuclei.
While most are multicellular, yeasts constitute a specialized group of unicellular fungi. Furthermore, some fungi are master shapeshifters; dimorphic fungi possess the unique capability to alternate between a unicellular yeast morphology and a multicellular mold morphology depending on environmental cues like temperature.
Fungal Reproduction
Fungi propagate through both asexual and sexual means:
- Unicellular yeast fungi typically reproduce asexually through an unequal cellular division process known as budding.
- Multicellular fungi frequently reproduce asexually by generating and releasing massive quantities of microscopic spores. These fungal spores are environmentally dispersed by forces such as wind, water currents, or animal vectors.
When fungi reproduce sexually, the process is uniquely decoupled into two stages:
- Plasmogamy: The physical fusion of the cytoplasm from two compatible fungal mycelia.
- Karyogamy: Hours, days, or even centuries later, the cells undergo the physical fusion of two haploid nuclei resulting in the formation of a single diploid nucleus.
Symbiotic Masterminds
Fungi do not exist in isolation. They form incredibly complex mutualistic relationships:
- Mycorrhizae characterize a mutually beneficial symbiotic association between actively growing plant roots and a network of fungal mycelia. The fungus extracts minerals from the soil for the plant, while the plant feeds the fungus sugars.
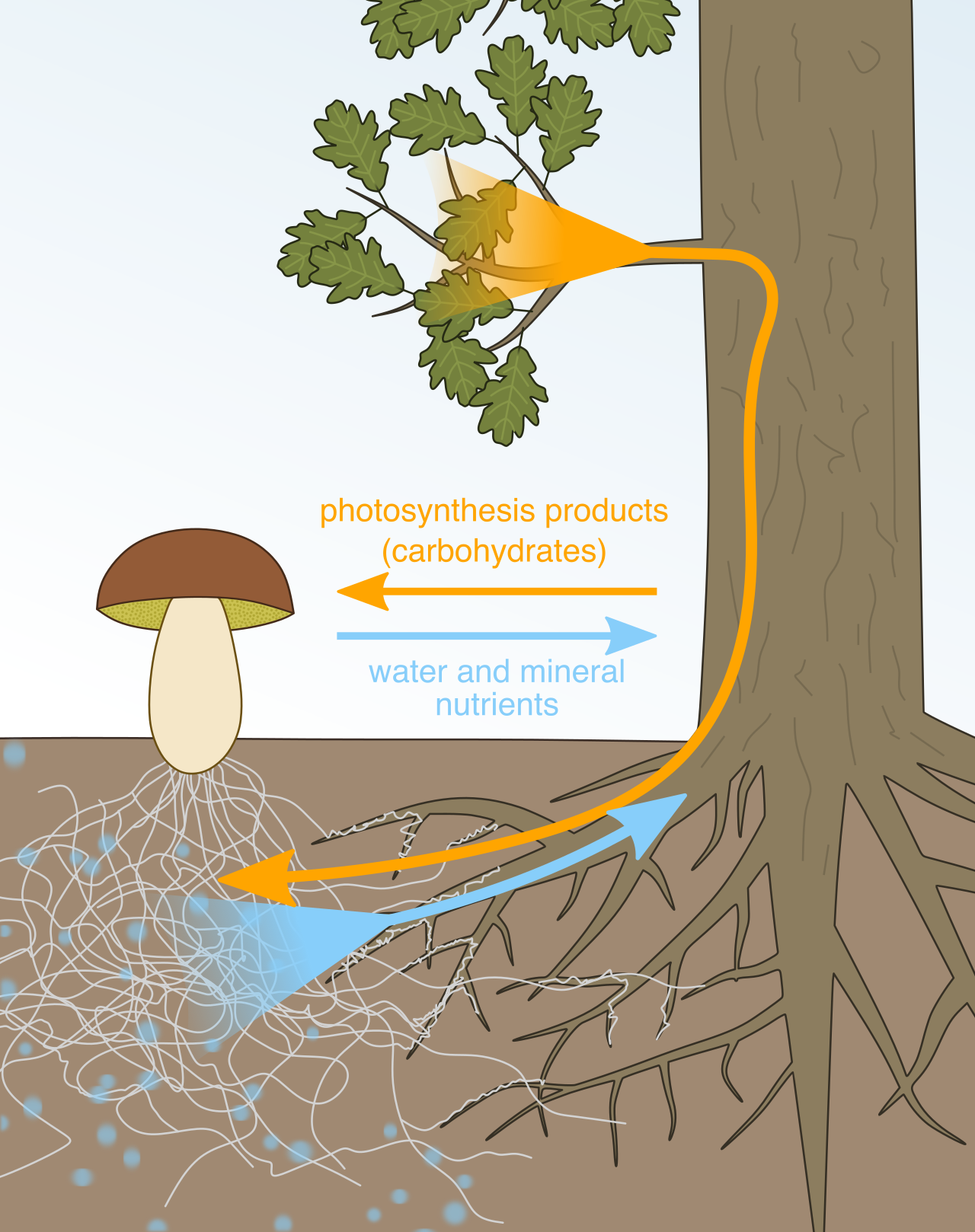
- Lichens constitute a composite organism emerging from a mutualistic symbiotic relationship between a fungal host and a photosynthetic partner (algae or cyanobacteria). They are pioneer organisms capable of breaking down bare rock into soil.
As a biology teacher, your goal is to help students see that this taxonomy is not a rigid list of rules, but the ongoing story of life's survival and adaptation. From the single-celled archaea boiling in a hydrothermal vent to the sprawling fungal mycelia beneath a forest floor, every organism operates under the same evolutionary pressures, utilizing elegant variations of the same cellular machinery.