Cell Cycle and Division
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Imagine attempting to transcribe an intricate, three-billion-letter manuscript entirely by hand without a single catastrophic error, and then physically partitioning your workshop to house two functional copies, all within a matter of hours. This staggering logistical feat—performed millions of times a second in the human body—is the eukaryotic cell cycle. It is not merely a sequence of biological events, but a masterclass in molecular quality control. Understanding how a single microscopic entity orchestrates its own duplication reveals the profound mechanisms that allow life to heal, grow, and perpetuate. It also exposes the devastating biological consequences when those precise mechanisms fail.
To teach the biology of cell division effectively, we must first dispel the illusion that cells are constantly dividing. In reality, the eukaryotic cell cycle consists of interphase and the mitotic phase, and it is characterized by meticulous preparation rather than perpetual splitting.
Interphase accounts for approximately 90% of the cell cycle duration in typical eukaryotic cells. Think of interphase not as a "resting" period—a common misconception among students—but as an era of intense, highly regulated biochemical activity.
Interphase is divided into three distinct phases called G1, S, and G2:
- G1 Phase (Gap 1): During the G1 phase, the cell grows physically larger. It is a period of bulk manufacturing. During the G1 phase, the cell synthesizes proteins necessary for normal cellular functions. It accumulates the metabolic energy and building blocks required for the momentous task ahead.
- S Phase (Synthesis): The S phase is the period of the cell cycle dedicated to DNA replication. This is the copying of the cellular manuscript. Crucially, DNA replication during the S phase forms two identical sister chromatids per chromosome. They remain tethered together, a vital structural requirement for their eventual segregation.
- G2 Phase (Gap 2): During the G2 phase, the cell synthesizes proteins specifically required for mitosis. The cell is stockpiling the exact biochemical machinery—like the tubulin needed for microtubules—that will drive the physical division of the nucleus.
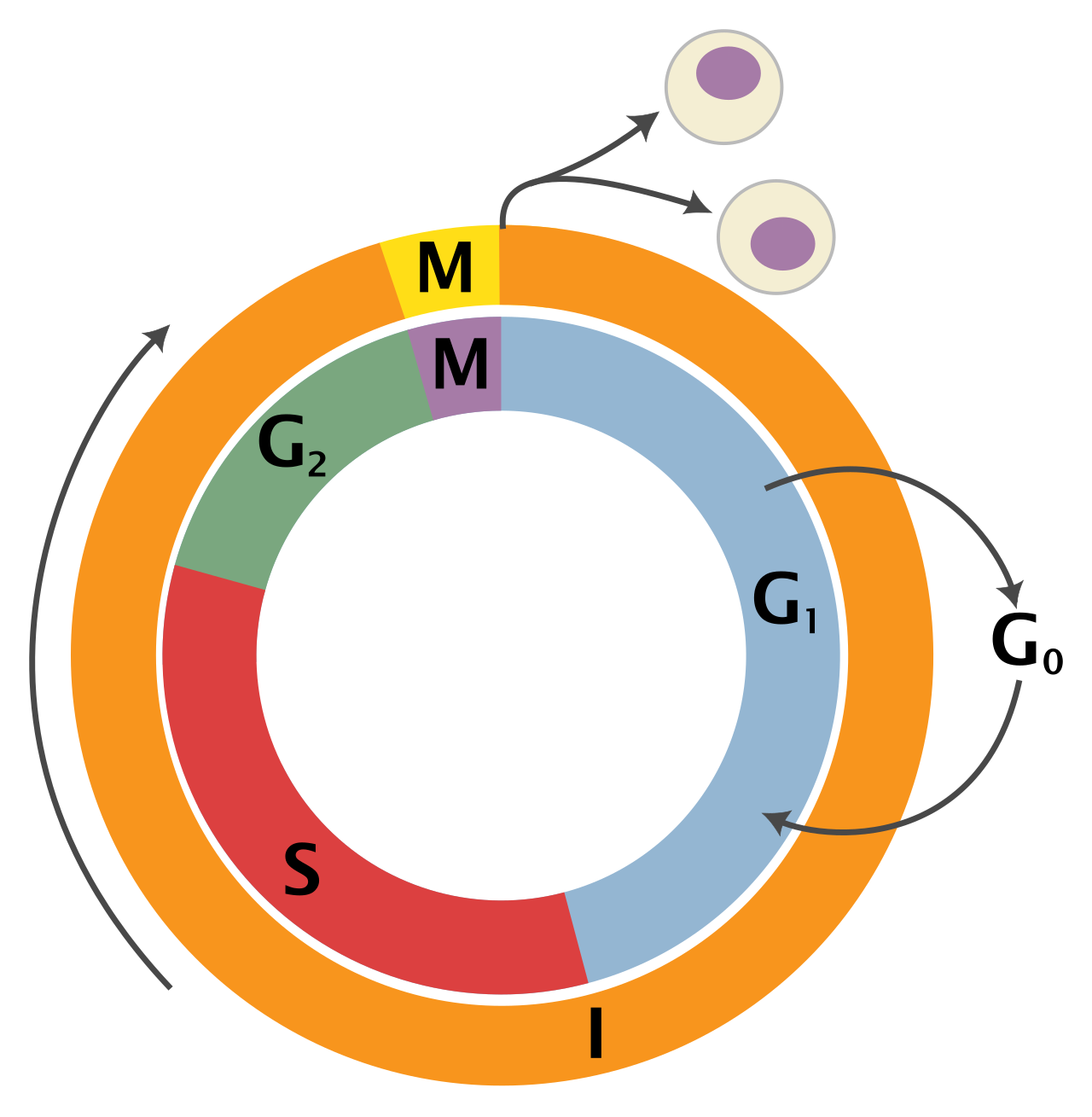
The G0 Phase: The Cellular Retirement
Not all cells are destined to divide. The G0 phase is a non-dividing cellular resting state. Cells may enter G0 temporarily due to environmental signals, but for many specialized tissues, it is a permanent exit from the cell cycle. For example, adult human nerve cells typically remain permanently in the G0 phase, as do adult human muscle cells. This biological reality explains why spinal cord injuries or heart attacks (which destroy cardiac muscle) are so devastating: the surrounding tissue generally cannot re-enter the cell cycle to replace the lost cells.
If interphase is the long preparation, the mitotic phase (M phase) is the dramatic climax. The M phase encompasses both the division of the genetic material and the division of the cell body.
Mitosis divides the eukaryotic cell nucleus into two genetically identical daughter nuclei. To achieve this elegant division without shredding the delicate DNA, the cell executes a highly choreographed dance. Mitosis comprises five sequential stages: prophase, prometaphase, metaphase, anaphase, and telophase.
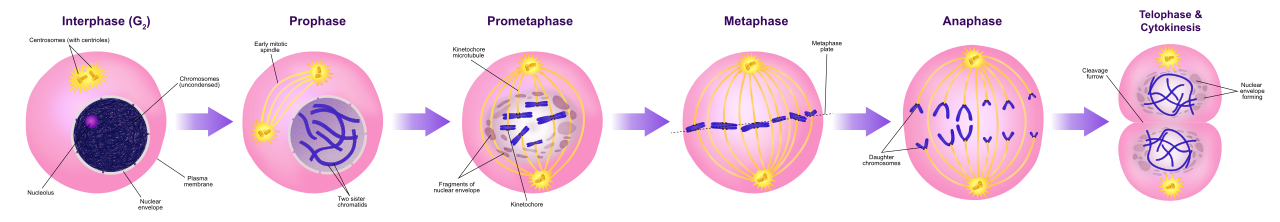
1. Prophase
During prophase, loosely packed chromatin condenses into distinct, visible chromosomes. This is akin to packing long, loose spools of thread into tight, movable bundles to prevent them from tangling when pulled apart. Simultaneously, the mitotic spindle begins to form during prophase, assembling from centrosomes at the edges of the cell.
2. Prometaphase
The physical barrier protecting the DNA must be removed. The nuclear envelope breaks down completely during prometaphase. With the chromosomes now exposed to the cytoplasm, spindle microtubules attach to specialized protein structures called kinetochores on the chromosomes. Think of kinetochores as molecular "handles" on the sister chromatids to which the microtubule "ropes" can latch.
3. Metaphase
A microscopic tug-of-war ensues until a state of perfect tension is reached. Chromosomes align along the cell equator during metaphase. They line up on what we call the metaphase plate, which is not a physical structure, but an imaginary plane located exactly between the two spindle poles.
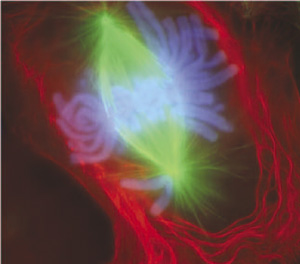
4. Anaphase
Once alignment is confirmed, the protein glue holding the chromatids together is cleaved. During anaphase, sister chromatids separate from each other. The mechanics of this are stunning: separated sister chromatids are pulled toward opposite spindle poles by shortening microtubules.
Crucial Conceptual Shift: Up until this exact moment, a chromosome consisted of two bound sister chromatids. However, separated sister chromatids are considered individual chromosomes during anaphase. A human cell entering anaphase briefly contains 92 individual chromosomes!
5. Telophase
As the identical sets of chromosomes arrive at opposite poles, the cell must reconstruct its normal interphase architecture. During telophase, a new nuclear envelope forms around each set of separated chromosomes at the cell poles. Because they no longer need to be moved, chromosomes begin to decondense back into chromatin during telophase, and the mitotic spindle breaks down completely.
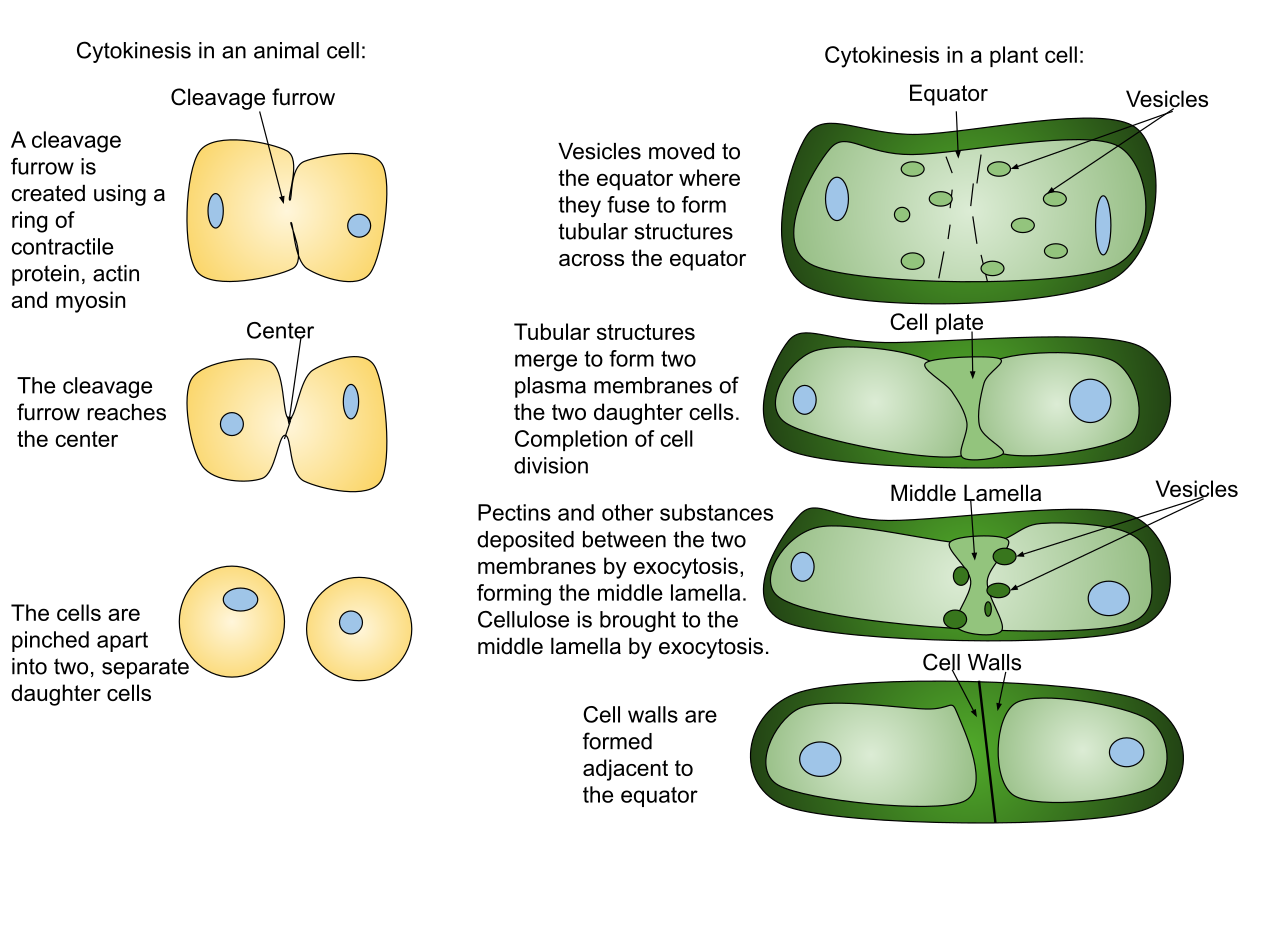
Cytokinesis: Cleaving the Cytoplasm
While telophase concludes the division of the nucleus, cytokinesis divides the cell cytoplasm to form two entirely separate daughter cells. Because of their distinct anatomies, animals and plants accomplish this quite differently:
| Cell Type | Cytokinesis Mechanism | The Process |
|---|---|---|
| Animal Cells | Cleavage Furrow | Animal cell cytokinesis occurs through the formation of a visible cleavage furrow. A biological "drawstring"—specifically, a contractile ring of actin microfilaments constricts to form the cleavage furrow in animal cells, pinching the cell in two. |
| Plant Cells | Cell Plate | Because plants possess rigid cell walls, they cannot be pinched. Plant cell cytokinesis occurs through the formation of a cell plate. To build this new partition from the inside out, Golgi-derived vesicles containing cell wall materials fuse at the cell equator to form the cell plate in plant cells. |
A cell cannot simply drift from one phase to the next; doing so would invite catastrophic errors, like trying to segregate DNA that hasn't finished copying.
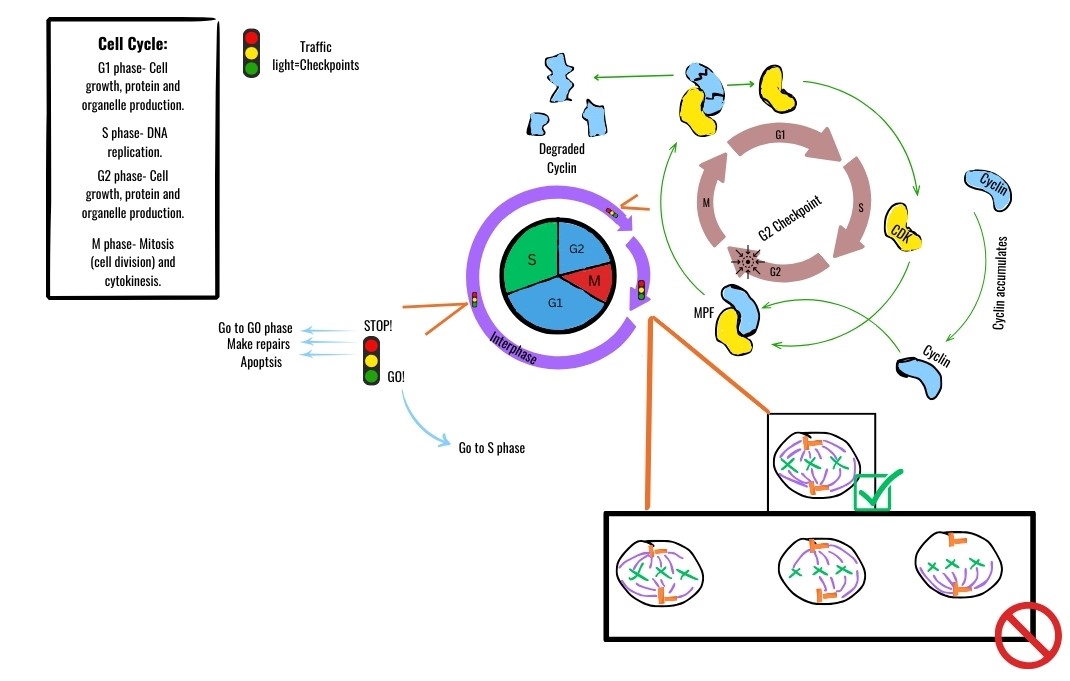
Cell cycle checkpoints are control mechanisms ensuring the proper sequential progression of the cell cycle.
There are three major checkpoints that act as rigorous quality-control inspectors:
- The G1 Checkpoint: This is the master switch. The G1 checkpoint determines whether a cell will proceed toward division or enter the non-dividing G0 phase. It assesses cell size, nutrient availability, and growth factors.
- The G2 Checkpoint: Occurring after DNA synthesis, the G2 checkpoint ensures that DNA replication is fully complete before mitosis is allowed to begin. Critically, the G2 checkpoint halts the cell cycle if unrepaired DNA damage is detected.
- The M Checkpoint: The M checkpoint occurs during the metaphase stage of mitosis. It is a physical tension sensor. The M checkpoint verifies that all chromosomes are properly attached to spindle microtubules before anaphase begins. If even one kinetochore is unattached, the cell waits. This prevents nondisjunction, which can lead to cells with abnormal chromosome numbers.
The Biochemical Engine: Cyclins and CDKs
The passage through these checkpoints is driven by an elegant biochemical engine consisting of two primary types of molecules:
- Cyclins: As their name implies, cyclin proteins fluctuate in concentration cyclically throughout the different phases of the cell cycle.
- Cyclin-dependent kinases (CDKs): Unlike cyclins, cyclin-dependent kinases are constant-concentration enzymes driving the cell cycle forward upon activation.
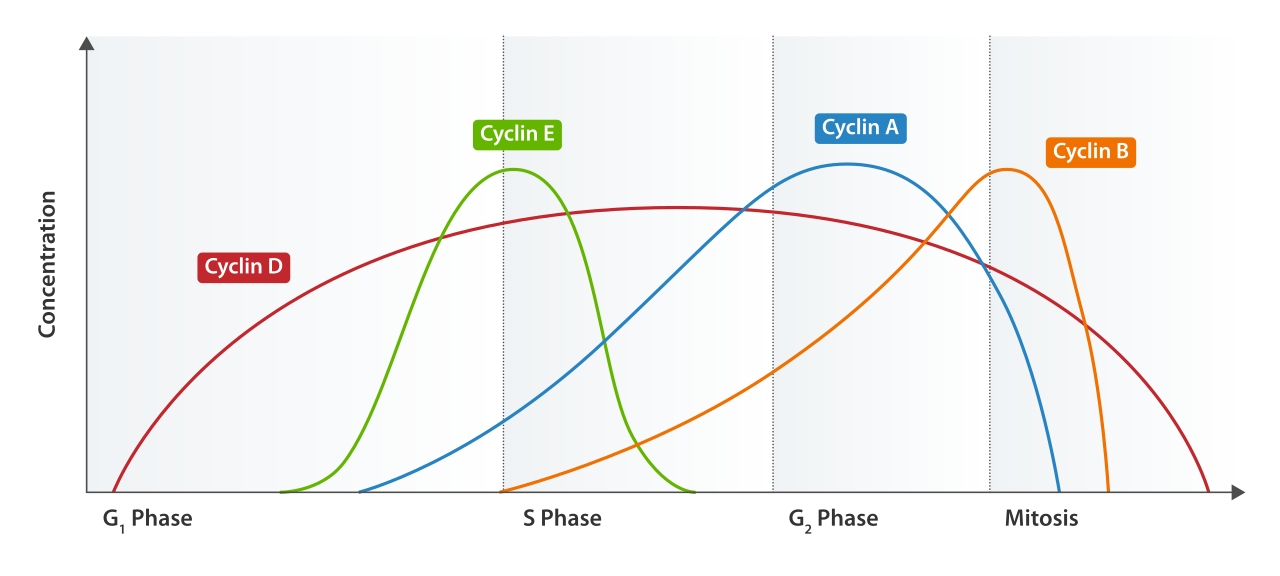
On their own, CDKs are inactive. Cyclin-dependent kinases require binding to specific cyclin proteins to become catalytically active. Once paired, active cyclin-CDK complexes phosphorylate target proteins to initiate specific downstream cell cycle events.
One of the most famous of these pairings is MPF. Maturation-promoting factor is a specific cyclin-CDK complex triggering passage past the G2 checkpoint into the M phase. MPF phosphorylates proteins that initiate nuclear envelope breakdown and chromosome condensation. As mitosis concludes, the cyclin portion of MPF is degraded, deactivating the CDK and resetting the system.
The strict regulation of the cell cycle is a matter of life and death. Cancer results directly from the loss of normal cell cycle control mechanisms. When cells ignore checkpoints, unregulated cell proliferation leads to the formation of abnormal cell masses called tumors.
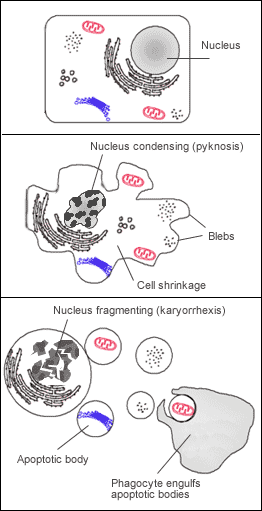
In a healthy organism, severe cellular damage triggers a failsafe. Apoptosis is a form of programmed cell death triggered by unrepairable DNA damage. It is cellular suicide for the greater good of the organism. Tragically, cancer cells frequently evade apoptosis despite possessing severe structural DNA damage, allowing them to survive and multiply their genetic errors.
The Genetics of Cancer: Oncogenes and Tumor Suppressors
To understand cancer functionally, you can use the analogy of a car's acceleration and braking systems. Cancer typically requires both the gas pedal to be stuck down and the brakes to be severed.
The "Gas Pedal" Genes:
- Proto-oncogenes are normal cellular genes stimulating typical cell growth and division. They are essential for life.
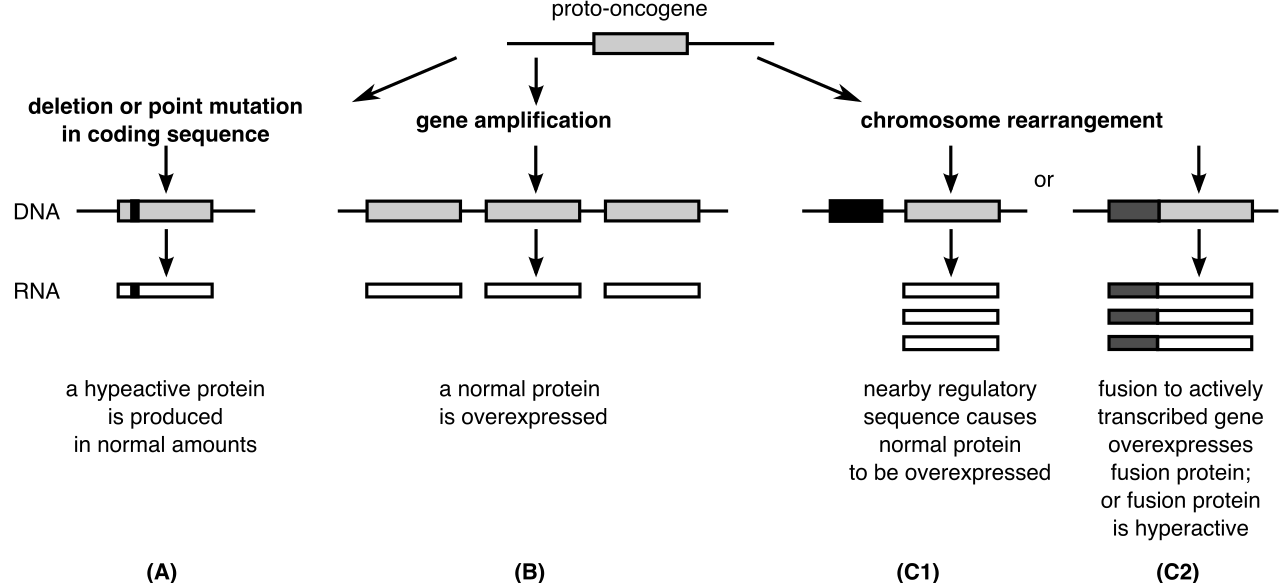
- However, when damaged by radiation, chemicals, or viral insertion, mutated proto-oncogenes become cancer-driving oncogenes.
- Once transformed, oncogenes drive continuous cell division even in the complete absence of external growth signals. The accelerator is permanently depressed.
The "Brake" Genes:
- Tumor suppressor genes encode proteins that inhibit cell division. They are the emergency brakes at the cell checkpoints.
- Furthermore, tumor suppressor genes encode proteins that actively promote apoptosis in defective cells.
- Because their function is protective, loss-of-function mutations in tumor suppressor genes remove vital inhibitory brakes on the cell cycle.
The Guardian of the Genome: The p53 Gene
If there is one specific gene every biology educator must know, it is p53. The p53 gene is a prominent tumor suppressor gene heavily involved in the G1 cell cycle checkpoint.
When DNA damage is detected, the p53 protein halts the cell cycle to allow adequate time for cellular DNA repair mechanisms. It acts as a molecular barricade. If the repair enzymes succeed, p53 releases the block. However, if the damage is catastrophic, the p53 protein forces the cell to initiate apoptosis if cellular DNA damage is deemed beyond repair.
Because of its critical position in cellular defense, losing p53 function is disastrous. Mutations in the p53 gene are found in more than half of all documented human cancers. Without p53, cells with fragmented or heavily mutated DNA divide recklessly, accumulating even more mutations with each cycle.
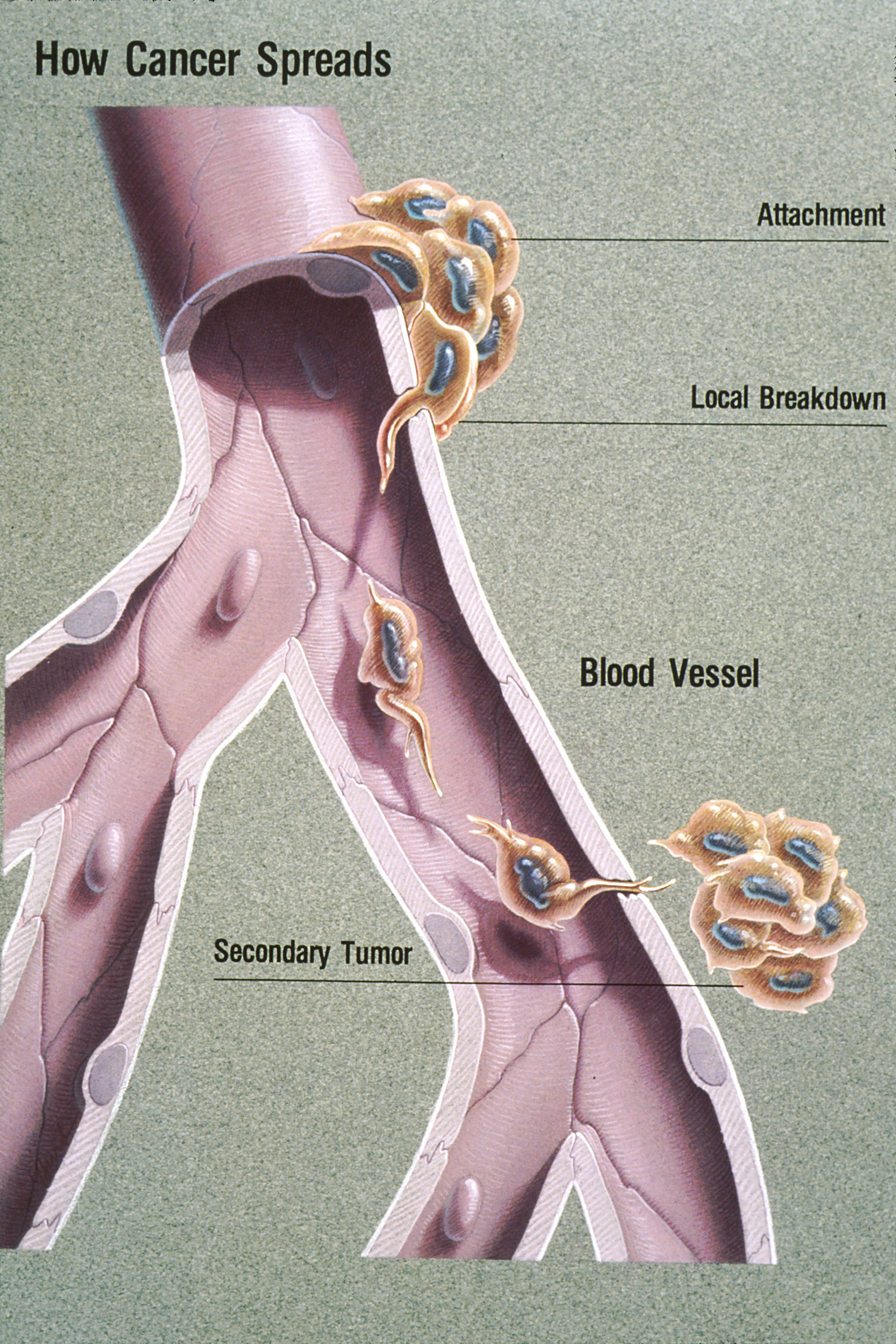
Malignancy and Metastasis
As a tumor grows and acquires more mutations, it becomes increasingly dangerous. While benign tumors remain localized, malignant tumors have the specific capacity to invade surrounding healthy tissues.
The ultimate, and most lethal, expression of this loss of control is metastasis. Metastasis is the active spread of cancer cells from the original primary tumor site to distant locations in the body, often traveling through the bloodstream or lymphatic system. A cell cycle defect that began as a microscopic malfunction in a single cell can ultimately overthrow the systemic homeostasis of the entire organism.