Cellular Respiration
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Energy bound within the covalent bonds of a glucose molecule is biologically inert. It is analogous to crude oil—highly energy-dense but fundamentally incompatible with the delicate, specialized nanomachinery of a living cell. Before the potential energy of organic macromolecules can drive the myriad endergonic reactions that sustain life, it must be refined into a universal biological currency. For a secondary biology educator, the pedagogical challenge lies in moving students past the rote memorization of chemical equations to a dynamic, mechanical understanding of bioenergetics. Mastering this topic requires revealing how biological molecules are systematically dismantled, how electrons physically cascade down microscopic gradients, and how the architecture of the cell is perfectly evolved to harvest this release of energy to maintain the highly ordered state we call life.
To teach cellular respiration effectively, you must first ensure your students understand the product being manufactured. ATP stands for adenosine triphosphate, the definitive energy carrier in biological systems.
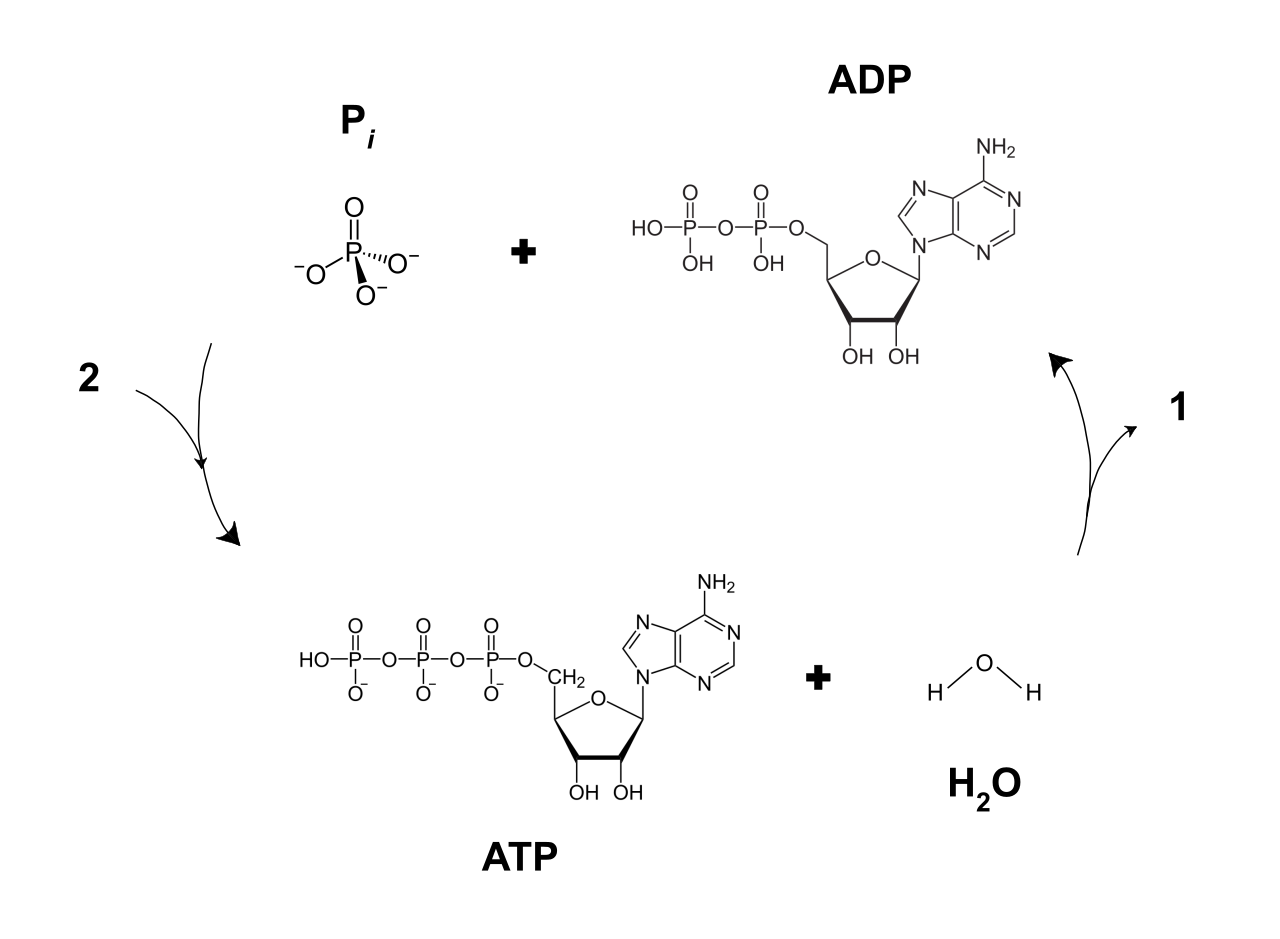
Structurally, adenosine triphosphate consists of an adenine nitrogenous base, a five-carbon ribose sugar, and a tail of three phosphate groups. The secret to its power lies in that phosphate tail. The covalent bonds between the phosphate groups of adenosine triphosphate store high potential energy due to electrostatic repulsion between the negatively charged oxygen atoms.
Teaching Analogy: Imagine holding three powerful, negatively charged magnets together. They desperately want to push apart. Compressing them together requires energy, and that mechanical energy is stored in the system. The phosphate groups in ATP act exactly like compressed springs.
When the cell needs to do work, it "releases the spring." The hydrolysis of the terminal phosphate bond in adenosine triphosphate releases energy to drive endergonic cellular work. This exergonic reaction—cleaving that outer phosphate—transfers energy to other molecules and produces adenosine diphosphate (ADP) and an inorganic phosphate molecule.
Because life requires a relentless input of energy, a cell's supply of ATP is depleted almost instantly. To survive, cells continuously regenerate adenosine triphosphate by adding an inorganic phosphate to adenosine diphosphate. Cellular respiration is simply the grand, multi-step process of powering that continuous regeneration.
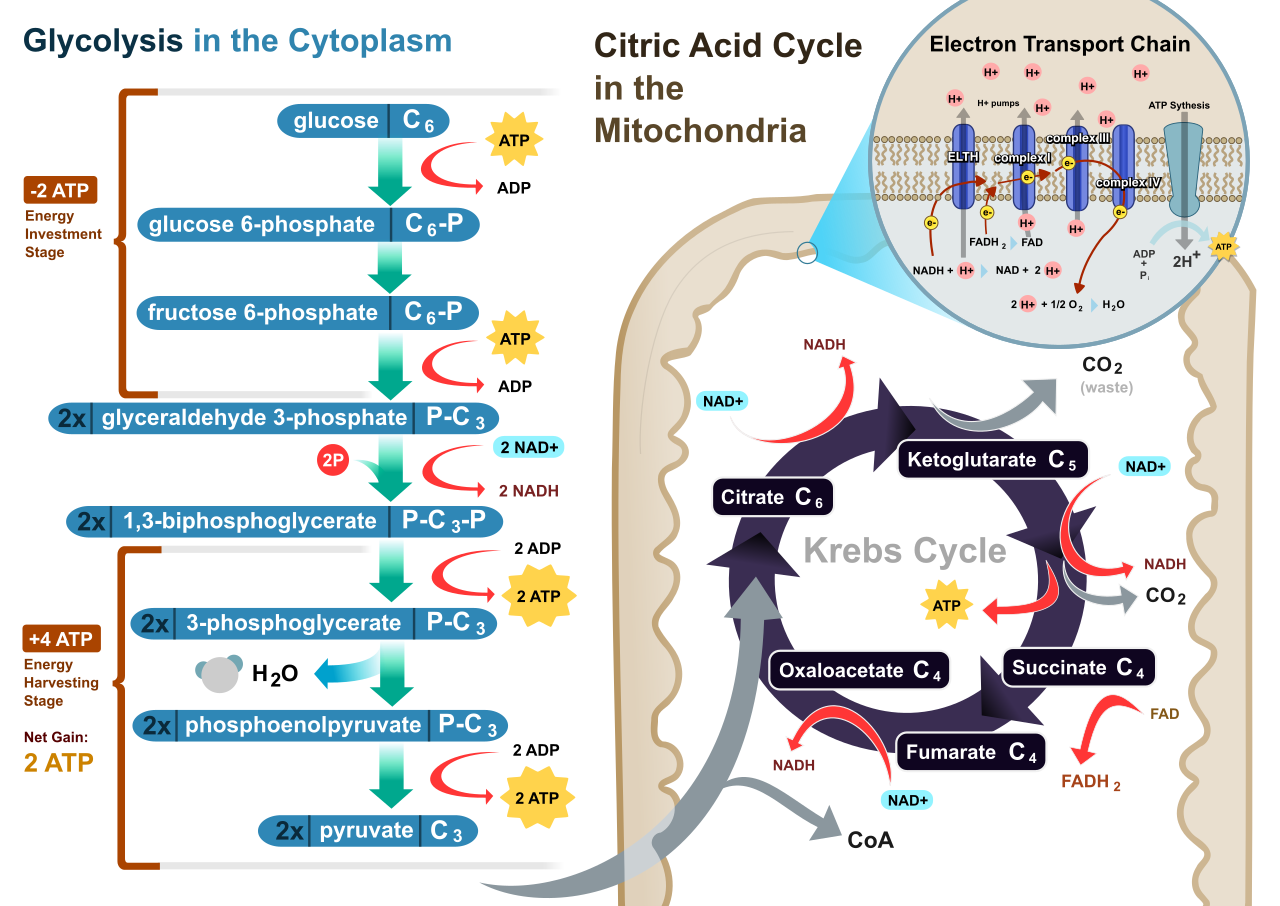
Glycolysis is the first biochemical stage of cellular respiration. From an evolutionary perspective, it is ancient. Glycolysis occurs in the cytoplasm of all living cells—from the bacteria in the soil to the neurons in your brain. Crucially, the biochemical pathway of glycolysis does not require oxygen to function, making it an entirely anaerobic primer for both aerobic and anaerobic systems.
Mechanically, glycolysis enzymatically cleaves one six-carbon glucose molecule into two three-carbon pyruvate molecules. This does not happen spontaneously; it requires an initial investment of cellular capital.
- The Energy Investment Phase: To destabilize the glucose molecule enough to split it, the initial energy investment phase of glycolysis consumes two adenosine triphosphate molecules per glucose molecule.
- The Energy Payoff Phase: As the sugar is broken apart and oxidized, the energy payoff phase of glycolysis produces four adenosine triphosphate molecules per glucose molecule.
- The Net Result: Because we invested two and gained four, glycolysis yields a net gain of two adenosine triphosphate molecules per glucose molecule.
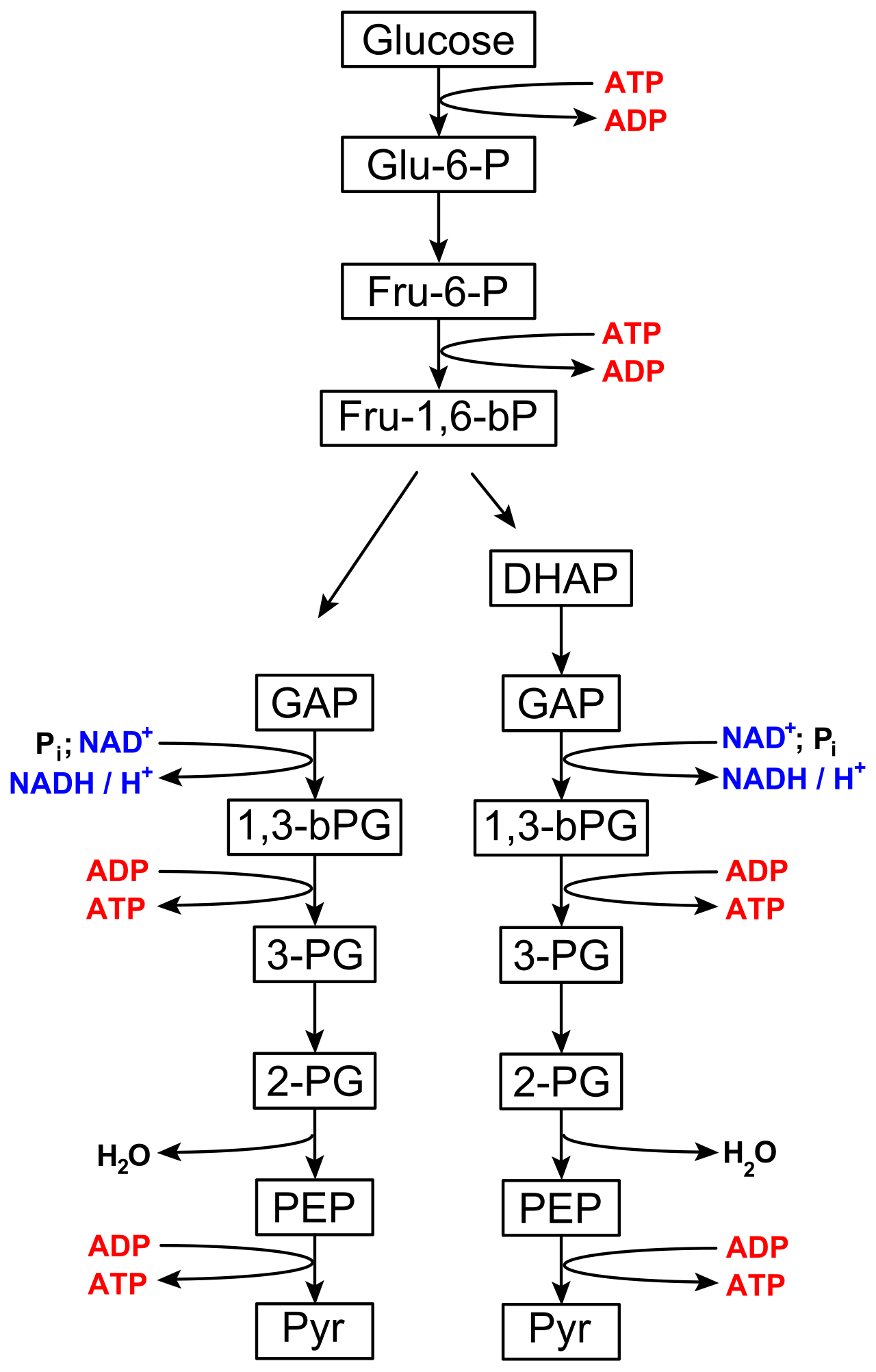
Adenosine triphosphate production during glycolysis occurs via substrate-level phosphorylation. This is a direct, enzymatic process where an enzyme strips a phosphate group from a metabolic substrate and snaps it directly onto ADP.
Glycolysis does not just produce ATP. It also extracts high-energy electrons. Over the course of the pathway, glycolysis produces two molecules of reduced nicotinamide adenine dinucleotide (NADH) per glucose molecule. NADH acts as an "electron shuttle bus," carrying highly reactive electrons to be cashed in later.
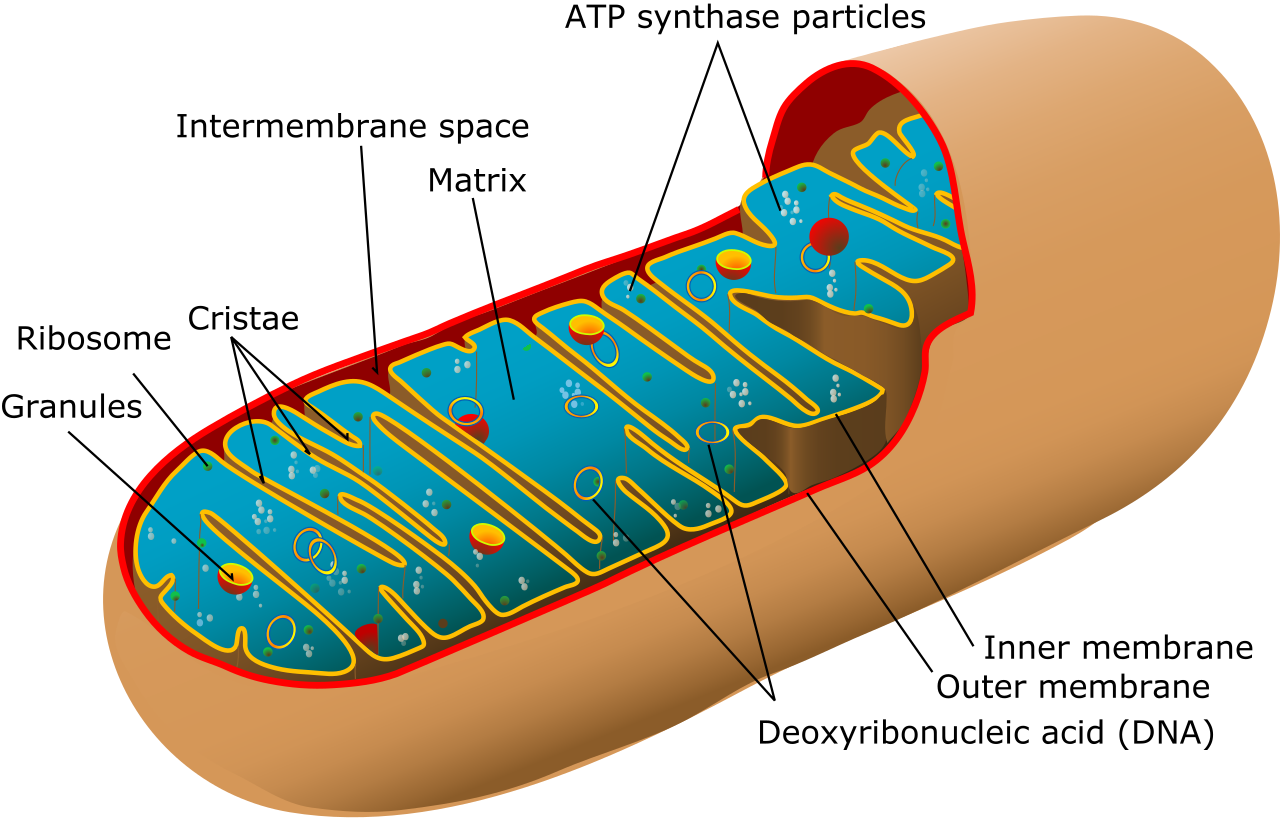
We now have two molecules of pyruvate sitting in the cytoplasm, still packed with potential energy. In eukaryotic cells, pyruvate molecules are actively transported from the cytoplasm into the mitochondrial matrix.
Once inside the mitochondria, a transition step occurs. During pyruvate oxidation, the three-carbon pyruvate molecule is decarboxylated to form a two-carbon acetyl group. This removal of carbon means that pyruvate oxidation releases one carbon dioxide molecule per pyruvate processed. (When your students ask where the CO2 they exhale comes from, this is the first place it is generated).
Simultaneously, the energetic electrons stripped during this breakdown are captured: pyruvate oxidation reduces one molecule of NAD+ to NADH per pyruvate processed. Finally, an enzyme attaches a coenzyme to the remaining fragment. Pyruvate oxidation binds the two-carbon acetyl group to Coenzyme A to form Acetyl-CoA. This molecule is now fully prepped to enter the cycle.
The citric acid cycle (also known as the Krebs cycle) takes place within the mitochondrial matrix in eukaryotic cells. It is a brilliant example of a biological loop.
The first step of the citric acid cycle combines a two-carbon acetyl group from Acetyl-CoA with the four-carbon oxaloacetate molecule. The combination of an acetyl group and oxaloacetate produces the six-carbon molecule citrate (hence the name of the cycle).
As citrate is enzymatically rearranged and oxidized, energy and electrons are systematically extracted. One complete turn of the citric acid cycle processes a single Acetyl-CoA molecule. (Since a single glucose yields two Acetyl-CoA molecules, the cycle turns twice per glucose).
The Yield of One Turn:
- Produces three NADH molecules.
- Produces one FADH2 molecule (another electron shuttle, slightly lower in energy than NADH).
- Produces one adenosine triphosphate molecule via substrate-level phosphorylation.
- Releases two molecules of carbon dioxide as metabolic waste.
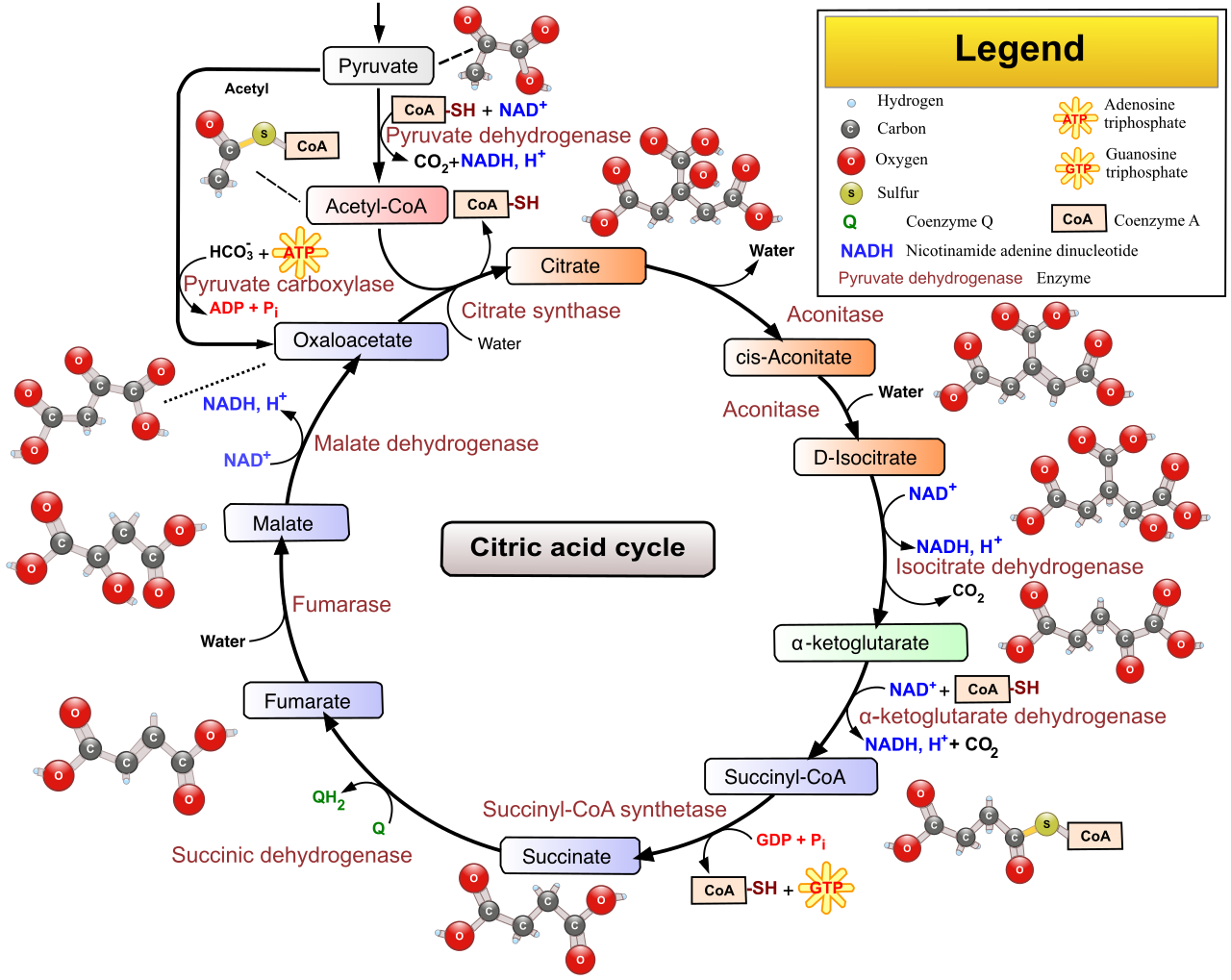
By the end of this cycle, the original glucose molecule is completely gone. The citric acid cycle completes the total metabolic breakdown of the original glucose molecule's carbon skeleton into carbon dioxide.
But why is it a cycle? Because the final enzymatic step of the citric acid cycle regenerates oxaloacetate at the end of the biochemical pathway to allow the cycle to continuously repeat.
Up to this point, we have only produced a measly 4 net ATP (2 from glycolysis, 2 from the citric acid cycle). But we have accumulated a massive stockpile of loaded electron shuttles: NADH and FADH2. Now, we cash them in.
Oxidative phosphorylation occurs at the inner mitochondrial membrane in eukaryotic cells. It is where the vast majority of cellular energy is generated. Oxidative phosphorylation consists of two coupled components: the electron transport chain and chemiosmosis.
The Electron Transport Chain (ETC)
NADH and FADH2 deliver high-energy electrons to the multiprotein complexes of the electron transport chain. As these electrons are passed from one protein to the next, they fall to lower and lower energy states. The sequential transfer of electrons through the protein complexes of the electron transport chain releases energy.
The cell does not let this energy dissipate as heat. Instead, the energy released by the electron transport chain is used to pump protons (H+ ions) from the mitochondrial matrix into the intermembrane space. The active pumping of protons creates a steep electrochemical gradient across the inner mitochondrial membrane. We have essentially pumped water up behind a dam.
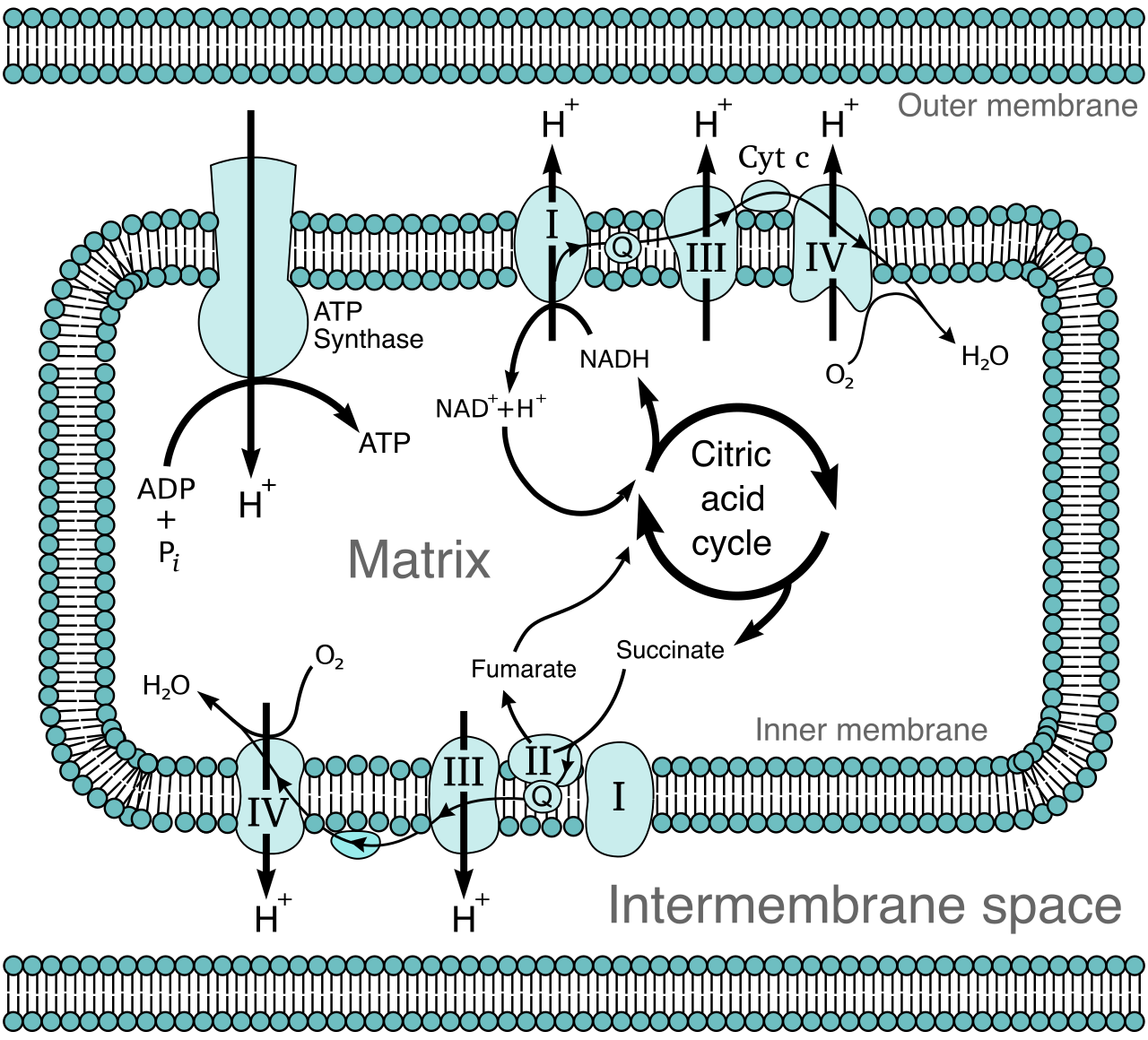
What happens to the electrons at the end of the chain? They must go somewhere, or the entire chain backs up. Oxygen functions as the final electron acceptor at the end of the aerobic electron transport chain. This is the molecular reason we breathe. The final electron acceptor oxygen combines with electrons and protons to form metabolic water.
Chemiosmosis
With a massive concentration of protons trapped in the intermembrane space, they desperately seek to diffuse back into the matrix. Chemiosmosis is the flow of protons down the electrochemical gradient back into the mitochondrial matrix.
However, the lipid bilayer is impermeable to protons. Protons flow back into the mitochondrial matrix exclusively through the membrane-bound enzyme ATP synthase. ATP synthase is quite literally a microscopic rotor. The exergonic flow of protons through ATP synthase provides the mechanical energy required to phosphorylate adenosine diphosphate into adenosine triphosphate.
The Final Tally
Thanks to this biological turbine, oxidative phosphorylation generates approximately 26 to 28 adenosine triphosphate molecules per glucose molecule in eukaryotes. When combined with glycolysis and the citric acid cycle, the theoretical maximum yield of aerobic cellular respiration is 36 to 38 adenosine triphosphate molecules per single glucose molecule. Aerobic respiration extracts significantly more usable chemical energy from a glucose molecule compared to fermentation pathways.
What happens when oxygen is unavailable to pull electrons through the ETC? The transport chain halts, NADH cannot unload its electrons, and without NAD+, even glycolysis shuts down. The cell will die without a backup plan.
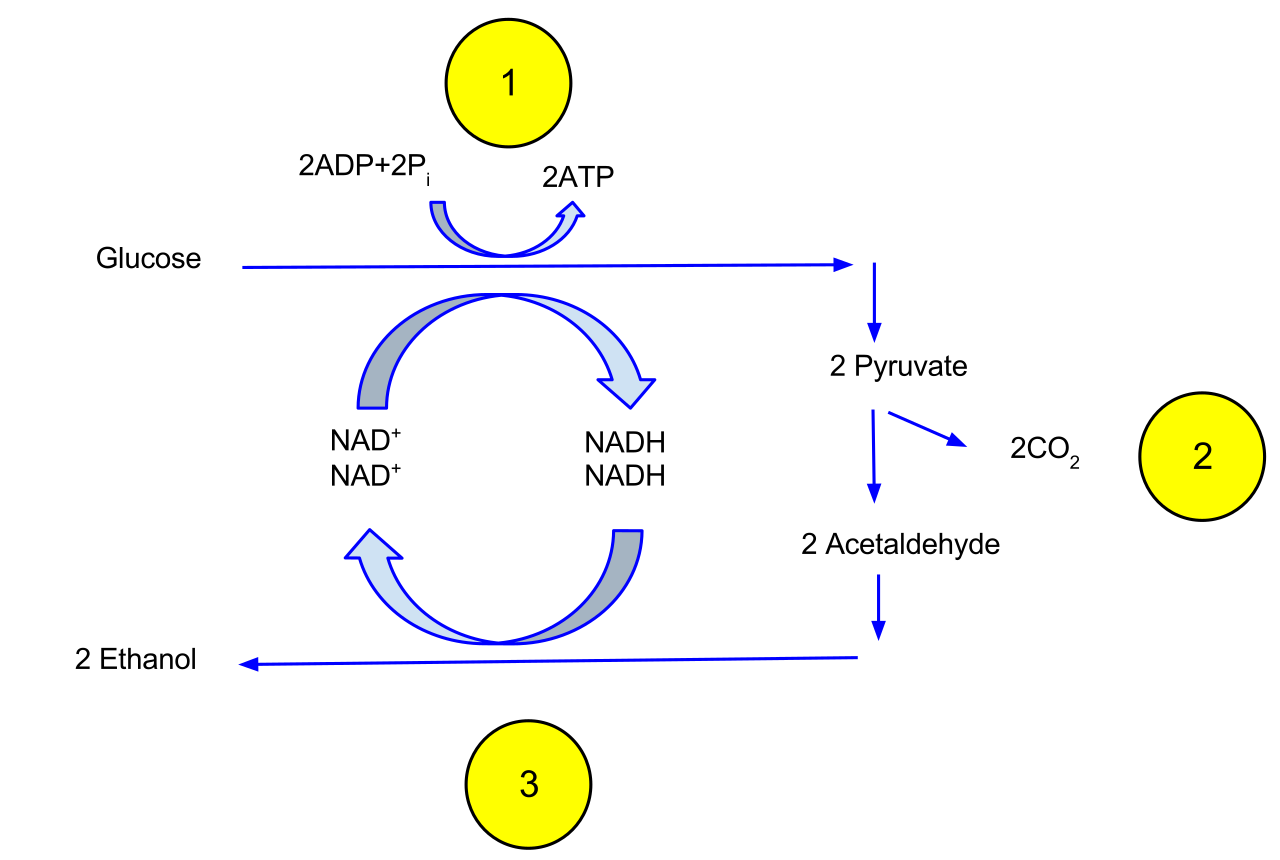
Fermentation allows glycolysis to continue producing adenosine triphosphate in the total absence of oxygen. Fermentation pathways do not utilize an electron transport chain, and fermentation pathways do not utilize the citric acid cycle. They rely exclusively on glycolysis.
The primary function of fermentation pathways is to oxidize NADH back into NAD+. The continuous supply of NAD+ regenerated by fermentation is required for the energy payoff phase of glycolysis to proceed. Without it, energy production flatlines. Because it relies entirely on glycolysis, fermentation produces a net yield of only two adenosine triphosphate molecules per glucose molecule.
There are two primary flavors of fermentation your students must know:
Lactic Acid Fermentation
Lactic acid fermentation enzymatically reduces pyruvate directly to lactate without releasing carbon dioxide. This process is highly relevant to human biology. Human skeletal muscle cells utilize lactic acid fermentation during periods of strenuous activity when cellular oxygen is depleted (like sprinting to catch a bus).
Alcoholic Fermentation
Alcoholic fermentation involves the decarboxylation of pyruvate to acetaldehyde. Because carbon is removed, alcoholic fermentation releases carbon dioxide gas as a metabolic byproduct (this is what makes bread rise!). The acetaldehyde intermediate in alcoholic fermentation is then reduced by NADH to form ethanol. Yeast cells perform alcoholic fermentation to generate adenosine triphosphate under anaerobic conditions.
Anaerobic Respiration vs. Fermentation
A common pedagogical pitfall is conflating fermentation with anaerobic respiration. They are not the same. Anaerobic respiration differs from fermentation because anaerobic respiration utilizes an electron transport chain.
How does it run a transport chain without oxygen? Anaerobic respiration employs a non-oxygen inorganic molecule as the final electron acceptor in the electron transport chain. These organisms (often extreme extremophilic bacteria) utilize different chemistry at the end of their ETC. Common final electron acceptors in anaerobic respiration include sulfate, nitrate, and elemental sulfur.
Summary Comparison for the Classroom
| Feature | Aerobic Respiration | Anaerobic Respiration | Fermentation |
|---|---|---|---|
| Needs Oxygen? | Yes | No | No |
| Uses an ETC? | Yes | Yes | No |
| Final Electron Acceptor | Oxygen | Inorganic molecule (Sulfate, Nitrate, Sulfur) | Organic molecule (Pyruvate or Acetaldehyde) |
| ATP Yield per Glucose | 36 - 38 ATP | Variable (less than aerobic, more than fermentation) | 2 ATP |
By anchoring these biochemical pathways in mechanical analogies—from compressed springs to hydroelectric dams—you equip your students to understand cellular respiration not as a list of trivial facts, but as the magnificent engine that drives the biosphere.