Enzyme Structure and Kinetics
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Imagine a living cell as a vast, densely packed metropolis where billions of precise chemical reactions must occur every second simply to keep the organism alive. If these reactions relied solely on the random, unassisted collisions of molecules at cellular temperatures, life would operate at a glacial pace, ultimately failing to sustain itself. This is the precise problem solved by enzymes, which are biological catalysts that speed up chemical reactions to the pace required for life. While the vast majority of biological enzymes are proteins, a vital evolutionary footnote exists in the form of ribozymes, which are RNA molecules capable of acting as biological catalysts. To understand how these molecular machines achieve their remarkable feats of acceleration, we must look beyond abstract chemical equations and examine the physical, three-dimensional geometry of the molecules themselves.
When you introduce enzymes to your future biology students, the most crucial concept to impart is that chemistry at the cellular level is a mechanical, tactile process. Molecules have specific shapes, and those shapes dictate their function.
A substrate is the specific reactant molecule upon which an enzyme acts. However, the substrate does not interact with the entire enzyme. Instead, it docks at the active site, which is the specific physical region on an enzyme where substrate molecules bind.
The unique three-dimensional shape of an enzyme active site determines enzyme specificity. This is not a generalized pocket; it is a highly specialized environment. The amino acid side chains within an active site create a highly specific chemical environment tailored precisely for one target substrate.
Historically, biologists used the lock-and-key model, which proposes that an enzyme active site and a substrate fit perfectly together without any structural changes. While useful as an introductory metaphor, the physical reality is far more dynamic. Modern enzymology relies on the induced fit model, which states that an enzyme active site undergoes a conformational change upon substrate binding. Think of it like a perfectly tailored baseball glove wrapping around a ball as it lands in the pocket. Crucially, the conformational change described by the induced fit model enhances the ability of the enzyme to catalyze the reaction by physically manipulating the substrate into a more reactive state.
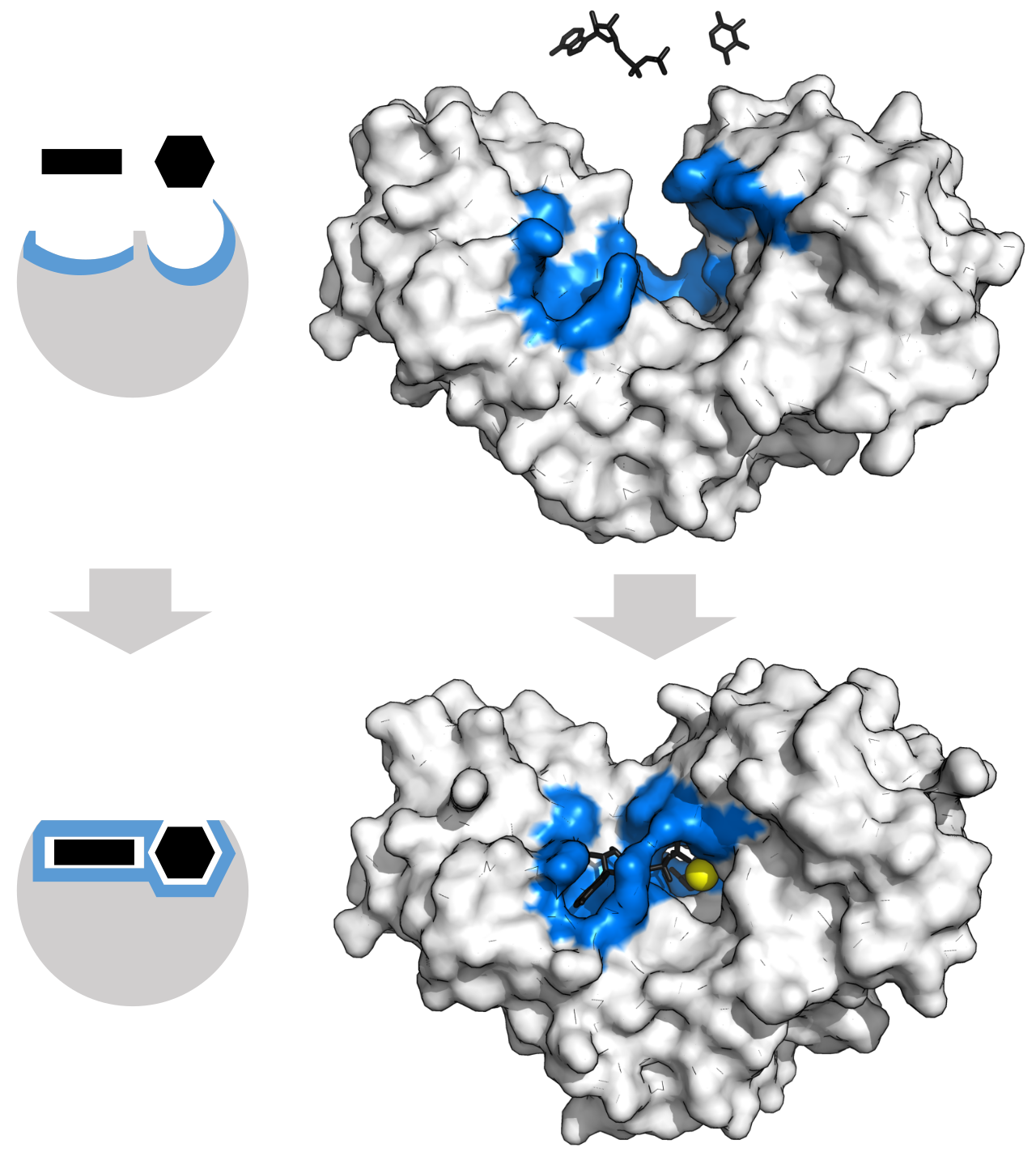
To understand how the induced fit accelerates a reaction, we must examine the energy landscape of the molecules involved.
Before a reactant can become a product, it must overcome an energy barrier. Activation energy is the initial energy input required to start a chemical reaction. This energy is needed to contort the reactant molecules into the transition state, which is an unstable, high-energy structural intermediate formed during a chemical reaction.
The Core Mechanism of Enzymes: Enzymes increase the rate of chemical reactions by lowering the activation energy barrier. They achieve this by physically stabilizing the transition state of the chemical reaction.
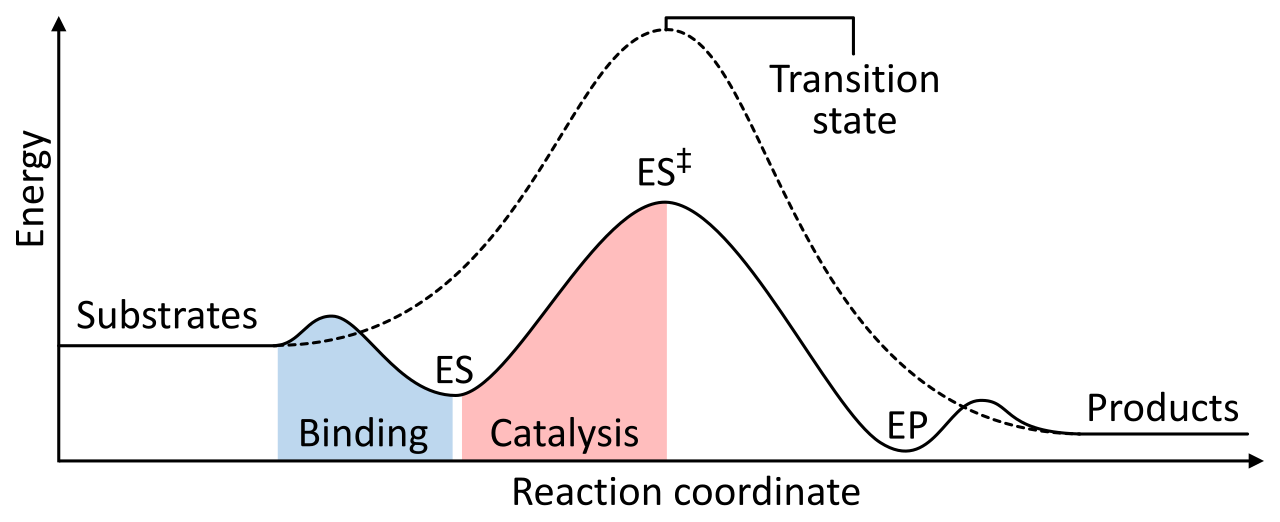
How do the amino acids in the active site actually do this?
- Enzymes can lower activation energy by bringing multiple substrates together in the correct spatial orientation. Instead of waiting for two molecules to randomly collide at the exact right angle, the enzyme physically holds them in perfect alignment.
- Enzymes can lower activation energy by providing a favorable microenvironment for the chemical reaction. For example, if a reaction requires an acidic environment to proceed, the active site may be lined with acidic amino acid side chains, creating a localized pocket of high reactivity.
What Enzymes Cannot Do
It is equally important for your students to understand the limits of catalysis. Gibbs free energy represents the usable energy in a chemical system available to do work.
While enzymes lower the energy required to start the reaction, enzymes do not change the overall Gibbs free energy difference between the reactants and the products. Because of this, enzymes do not change whether a chemical reaction is exergonic (energy-releasing) or endergonic (energy-consuming). Furthermore, in reactions that can go forward and backward, enzymes do not alter the final equilibrium point of a reversible chemical reaction. They simply help the system reach that inevitable equilibrium much faster.
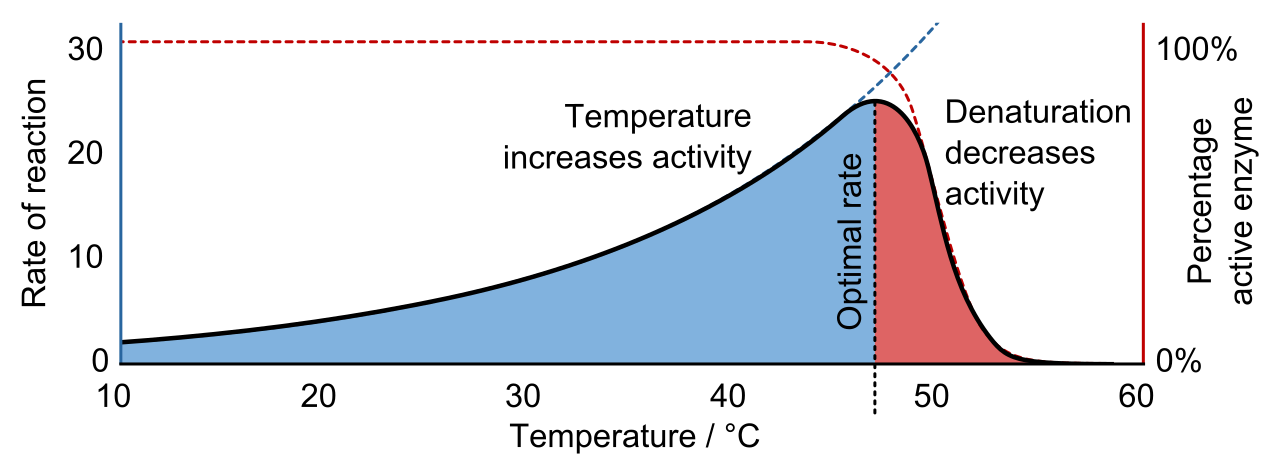
Because an enzyme's function relies entirely on its delicate three-dimensional folding, any environmental factor that disrupts this shape will destroy its catalytic ability.
The rate of an enzyme-catalyzed reaction generally increases with higher temperatures until an optimal temperature is reached. Heat means kinetic energy; molecules move faster, and substrates collide with active sites more frequently. However, there is a tipping point. High temperatures beyond the optimal point disrupt hydrogen bonds and noncovalent interactions within an enzyme molecule.
When these fragile bonds break, the protein begins to unfold. Denaturation is the loss of the functional three-dimensional structure of a protein. While slight unfolding might be reversible, extreme heat causes irreversible enzyme denaturation. A denatured enzyme loses catalytic function due to the destruction of the active site architecture. You can no longer catalyze a reaction if the precise microenvironment of the active site has melted into a tangled ribbon of amino acids.
Similarly, enzymes are exquisitely sensitive to acidity. Each specific enzyme functions optimally within a narrow pH range. Deviations from an optimal pH alter the ionization states of amino acid side chains within an enzyme. If an amino acid that requires a positive charge suddenly loses it due to a pH shift, extreme pH changes disrupt the ionic bonds stabilizing the tertiary structure of an enzyme. Consequently, extreme environmental pH changes cause enzyme denaturation.
Nature, however, adapts enzyme structures to their specific environments. For instance, pepsin is a human digestive enzyme that functions optimally at a highly acidic pH of approximately 2, perfectly suited for the harsh, hydrochloric acid-filled environment of the human stomach.
Proteins are incredibly versatile, but sometimes the standard 20 amino acids aren't enough to perform complex chemistry, such as shuttling electrons. For this, enzymes rely on accessories.
Cofactors are non-protein chemical compounds required for the biological activity of specific enzymes.
- Inorganic cofactors: Zinc, iron, and copper ions frequently act as inorganic cofactors for enzymes. This is why these trace minerals are essential in the human diet.
- Organic cofactors: Coenzymes are organic cofactors required for proper enzyme function. When your students ask why they need to eat their vitamins, you can give them a literal molecular answer: many human vitamins function directly as coenzymes or as precursors to essential coenzymes. Without vitamins, the enzymes cannot assume their catalytic shape, and vital metabolic pathways grind to a halt.
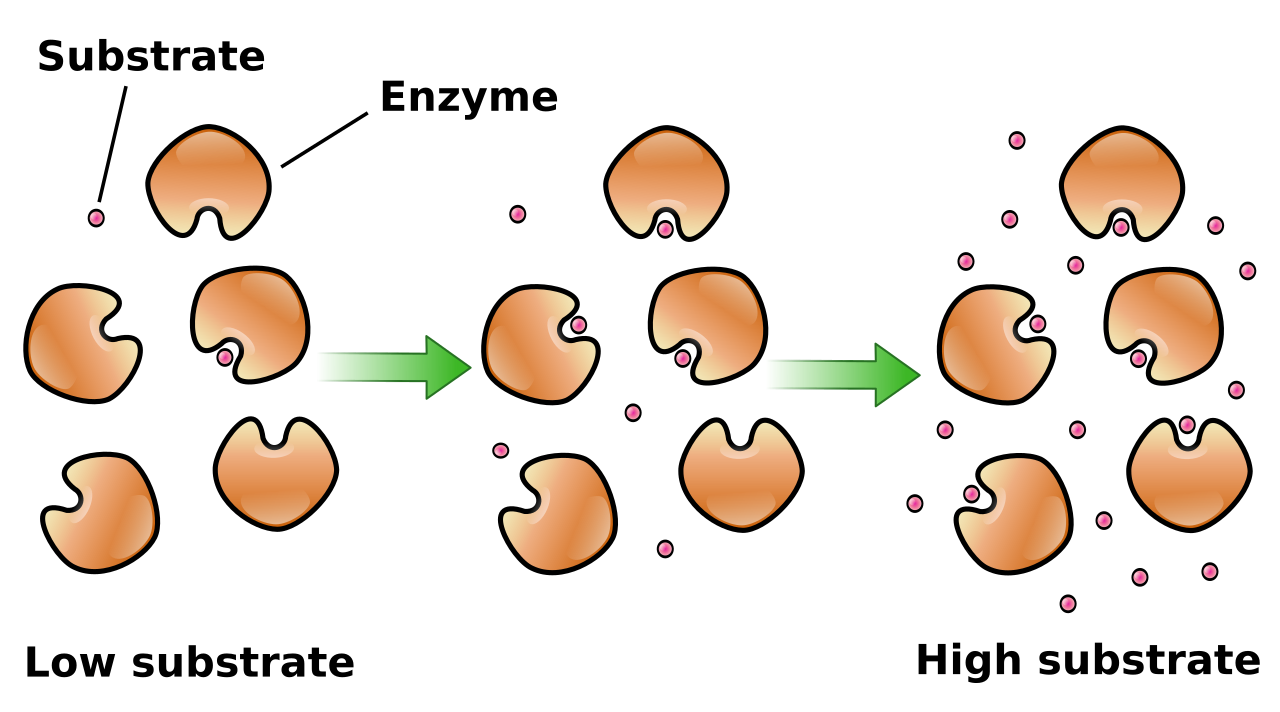
To analyze how effectively an enzyme works, biochemists use specific kinetic parameters. When analyzing reaction rate data, two numbers tell the story of the enzyme's efficiency:
- Maximum velocity (Vmax) is the peak rate of an enzyme-catalyzed reaction when all enzyme active sites are saturated with substrate. At this point, adding more substrate will not speed up the reaction, because every available enzyme is already working at top speed.
- The Michaelis constant (Km) is the exact substrate concentration at which the reaction rate is exactly half of the maximum velocity.
Km is an inverse measure of affinity. A low Michaelis constant value indicates a high affinity of an enzyme for the specific substrate. If Km is low, it means the enzyme requires very little substrate to reach half its maximum speed, because it is incredibly effective at grabbing and binding the substrate molecules floating by.
A cell cannot simply leave all its enzymes running at maximum velocity; metabolism requires exquisite regulation. Cells use various inhibitory molecules to act as the "brakes" on metabolic pathways.
Competitive vs. Noncompetitive Inhibition
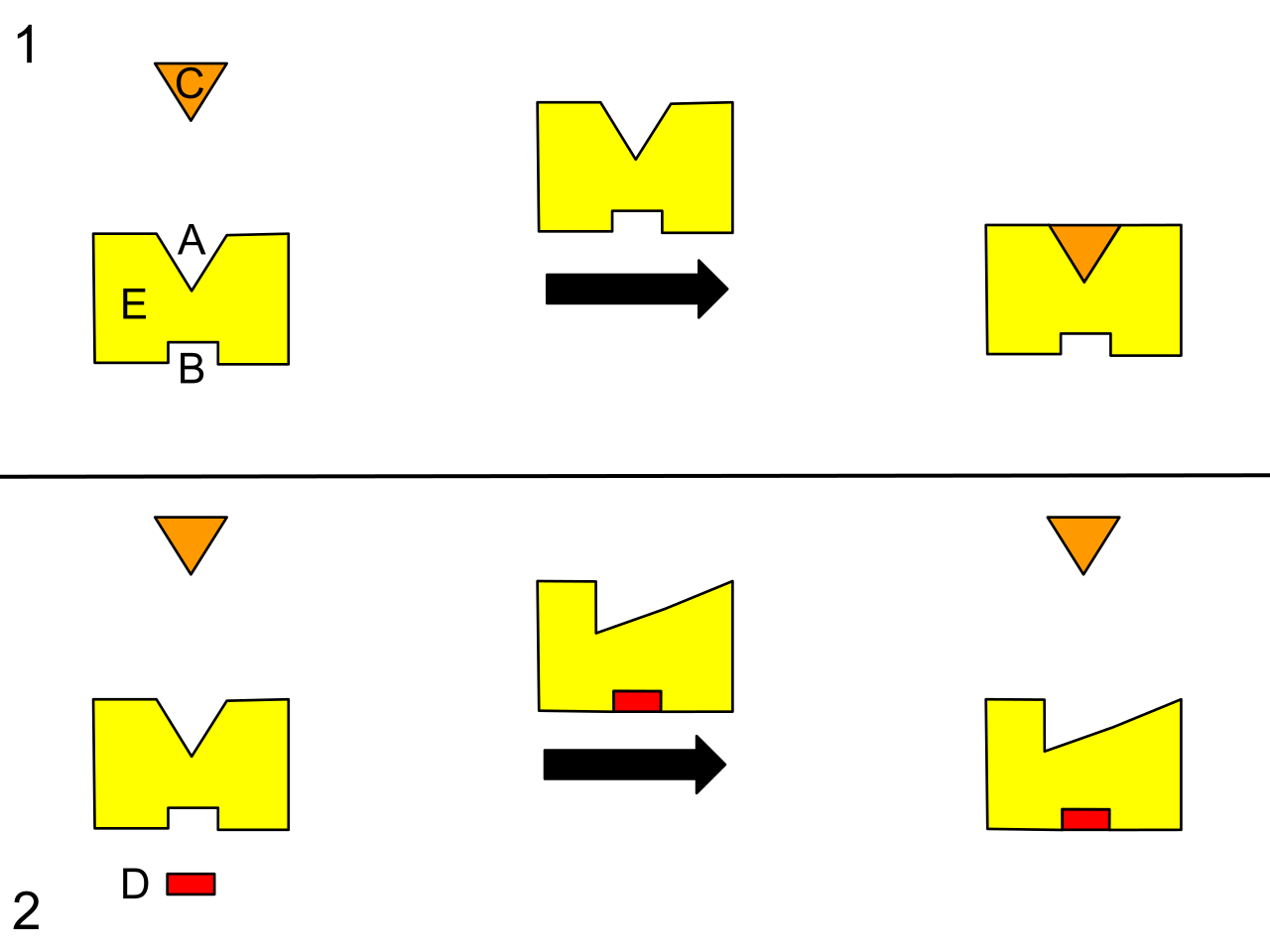
Competitive inhibitors are molecules that structurally resemble the normal substrate of an enzyme. Because of this molecular mimicry, competitive inhibitors bind directly to the active site of a target enzyme. By sitting in the active site, competitive inhibitors physically block the normal substrates from entering.
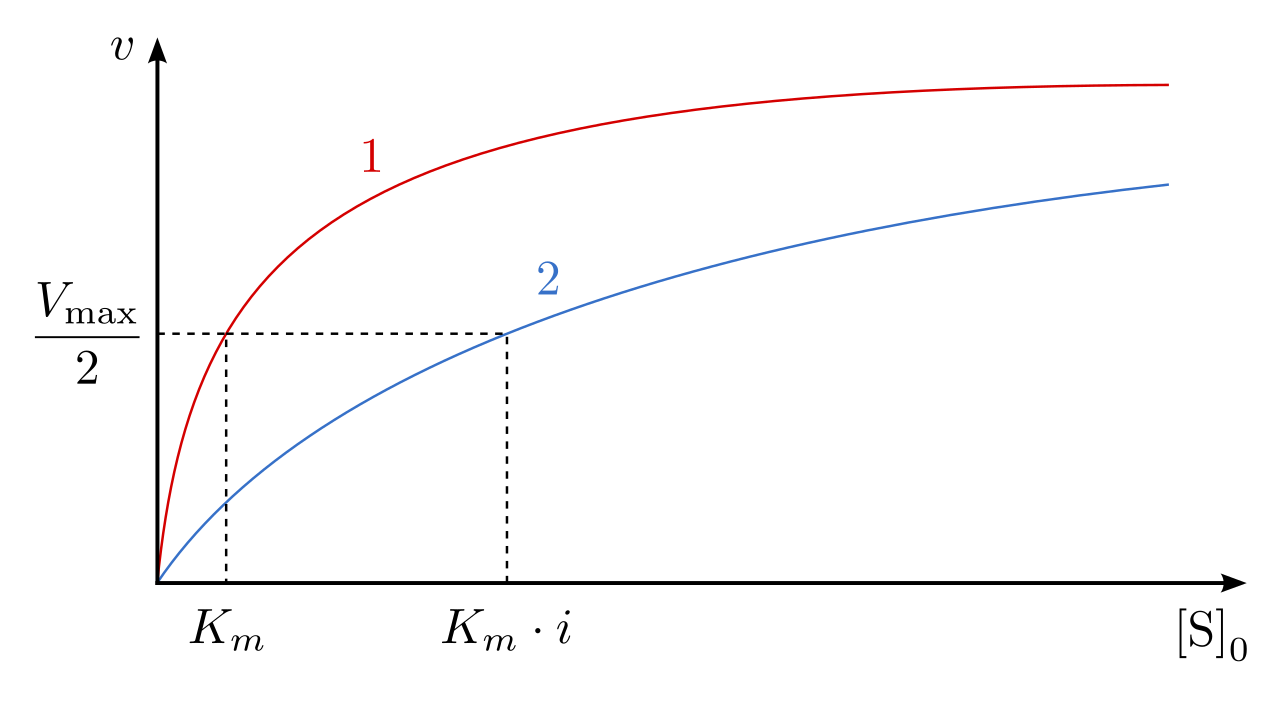
- The kinetic result: Because the inhibitor and substrate are competing for the exact same physical space, the inhibitory effect of a competitive inhibitor can be overcome by increasing the substrate concentration in the surrounding solution. If you flood the system with substrate, the substrate will outcompete the inhibitor.
- Mathematical impact: Because it takes more substrate to reach the same rate of reaction, a competitive inhibitor increases the apparent Michaelis constant (Km) of an enzyme-catalyzed reaction. However, a competitive inhibitor does not change the maximum velocity (Vmax) of an enzyme-catalyzed reaction; if you add enough substrate, the enzyme can still eventually reach its absolute top speed.
Noncompetitive inhibitors, on the other hand, play by different rules. Noncompetitive inhibitors bind to an allosteric site rather than the active site of an enzyme. An allosteric site is a receptor region on an enzyme distinct from the active site.
- The physical result: The binding of a noncompetitive inhibitor causes a conformational change in the overall shape of the enzyme. Crucially, the conformational change caused by a noncompetitive inhibitor alters the shape of the active site. The altered active site shape caused by a noncompetitive inhibitor prevents the enzyme from effectively catalyzing the reaction.
- The kinetic result: Because the inhibitor binds to a completely different site, increasing substrate concentration does not overcome the effects of a noncompetitive inhibitor. The active site is structurally broken, no matter how much substrate is available.
- Mathematical impact: Because a portion of the enzyme pool is functionally removed from the system, a noncompetitive inhibitor decreases the maximum velocity (Vmax) of an enzyme-catalyzed reaction. However, a noncompetitive inhibitor does not change the apparent Michaelis constant (Km) of an enzyme-catalyzed reaction, because the functional enzymes remaining in the pool still bind the substrate with the same native affinity.
| Inhibitor Type | Binding Location | Can be overcome by more substrate? | Effect on Vmax | Effect on Apparent Km |
|---|---|---|---|---|
| Competitive | Active Site | Yes | No change | Increases |
| Noncompetitive | Allosteric Site | No | Decreases | No change |
Allosteric Regulation and Feedback Inhibition
The concept of allosteric binding isn't just for toxins or external inhibitors; it is a vital internal mechanism. Allosteric regulation involves the binding of a regulatory molecule to an enzyme at a site distinct from the active site. This allosteric regulation alters the functional properties and catalytic activity of an enzyme, either turning it "off" (inhibition) or turning it "on" (activation).
This mechanism elegantly leads to feedback inhibition, which occurs when the end product of a metabolic pathway acts as an inhibitor for an enzyme earlier in the same pathway. Imagine a factory assembly line: when the warehouse is full of the final product, that product itself presses the "stop" button on the very first conveyor belt. This prevents the cell from wasting valuable chemical energy and resources synthesizing molecules it already has in abundance.
By mastering the physical relationships between active sites, activation energy, and kinetic variables, you provide your students with the foundation they need to understand metabolism not as a list of abstract equations to memorize, but as a dynamic, deeply logical, and tightly regulated molecular machinery.