DNA Replication and Transcription
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Imagine a central school district office holding the singular, irreplaceable architectural blueprint for every building in the region. If a contractor needs to build a new library, the district never sends the original, delicate blueprint to a muddy construction site. Instead, a clerk makes a temporary, working photocopy and sends that out to the bricklayers. In molecular biology, the original blueprint is DNA, the photocopy is RNA, and the bricklayers are the ribosomes assembling the building: a protein. This directional flow of information is the absolute foundation of all living systems. As an educator, your ability to make these invisible molecular mechanics intuitive for your students is what will bridge the gap between abstract biochemistry and the vibrant, macroscopic diversity of life they observe every day.
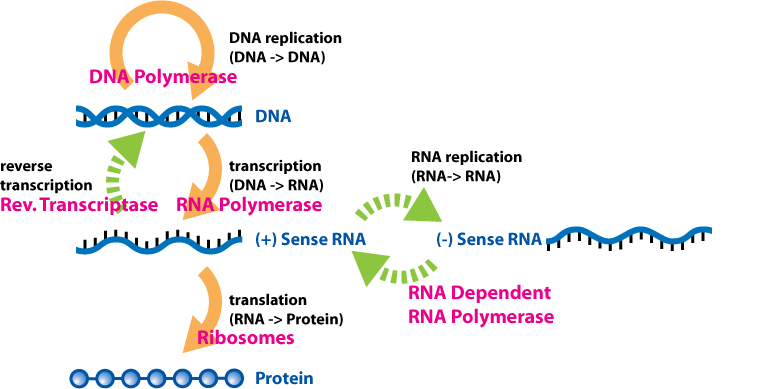
The overarching framework governing this information transfer is known as the central dogma of molecular biology. Francis Crick first articulated the central dogma of molecular biology in 1958, establishing that the flow of genetic information strictly moves from DNA to RNA to protein.
To teach this effectively, break it down into two distinct, sequential operations:
- Transcription is the process that transfers genetic information from a DNA sequence to an RNA molecule.
- Translation is the process that uses the sequence of an mRNA molecule to direct the synthesis of a polypeptide.
Pedagogical Note: A common student misconception is that the central dogma is an unbreakable law. It is not. You must introduce them to reverse transcription, a process that transfers genetic information from RNA back into DNA. Retroviruses, such as HIV, utilize reverse transcription to convert their viral RNA genomes into DNA, which is then integrated into the host's genome. This mechanism is a spectacular bridge between molecular genetics and evolutionary biology, showing how viruses hijack cellular machinery to survive.

Before a cell can divide, it must ensure the new cell receives a perfect copy of the genetic blueprint. DNA replication is an exercise in high-fidelity mass production.
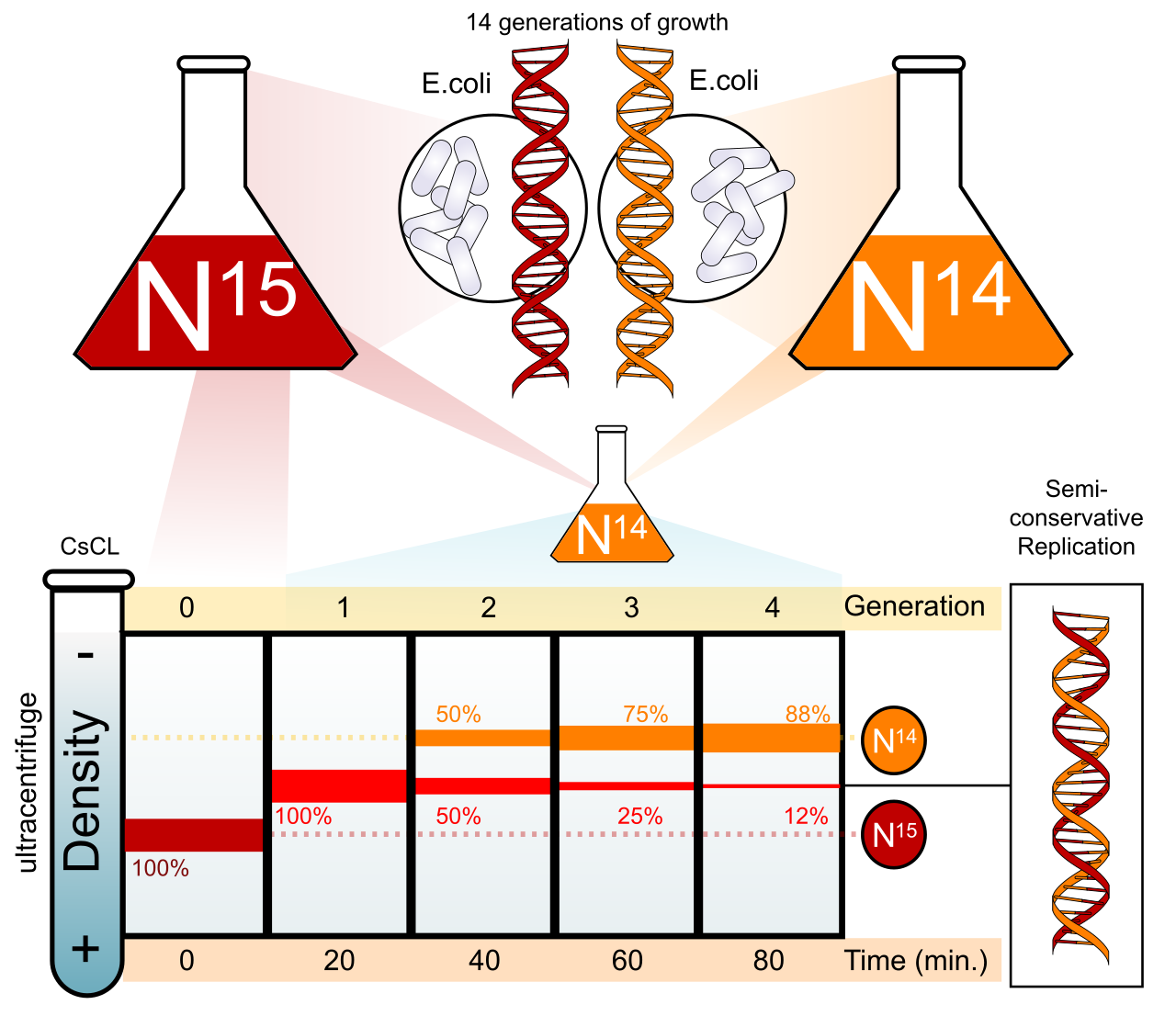
The Semi-Conservative Model
DNA replication follows a semi-conservative model. In semi-conservative replication, each newly synthesized DNA double helix contains one original parent strand and one completely new daughter strand.
When your students ask how we actually know this, introduce them to the Meselson-Stahl experiment, which provided the experimental proof for the semi-conservative model of DNA replication. By growing bacteria in heavy nitrogen (15N) and transferring them to light nitrogen (14N), Meselson and Stahl used density gradient centrifugation to show that after one generation, all DNA was uniformly of intermediate density—perfectly disproving both conservative and dispersive models. This is a masterclass in the Science and Engineering Practice of interpreting experimental data.
The Mechanics of the Replication Fork
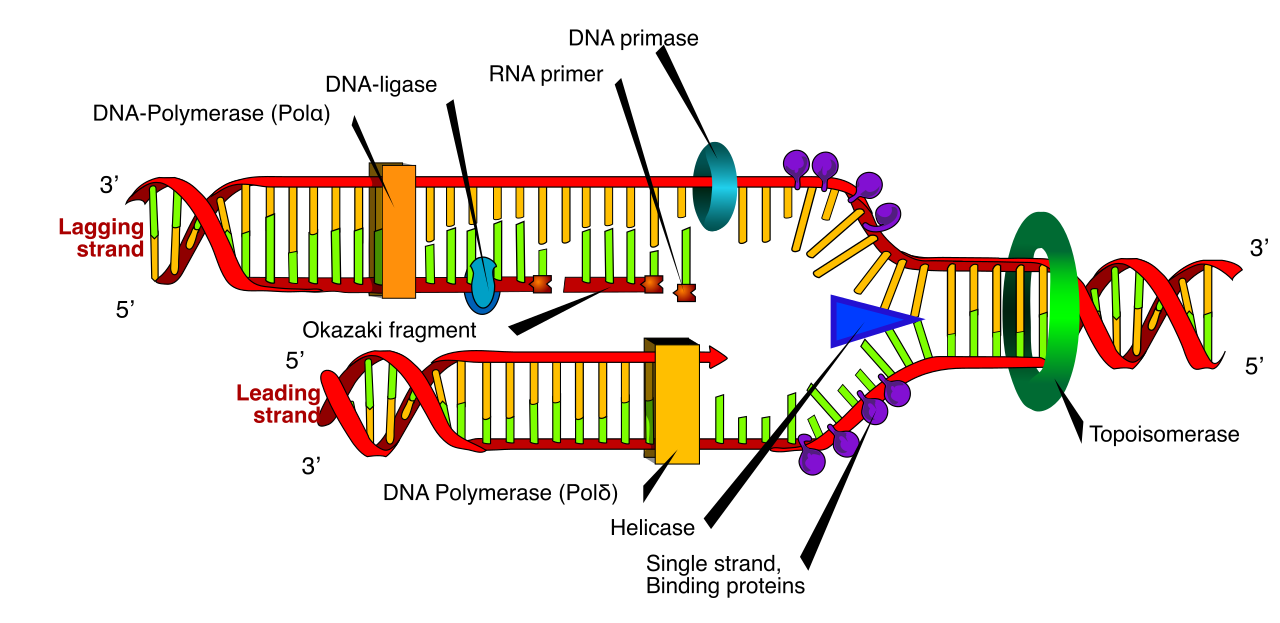
The complexity of DNA replication stems from one strict geometric rule: DNA synthesis always proceeds in the 5-prime to 3-prime direction. This happens because DNA polymerases can only add new nucleotides to the free 3-prime hydroxyl group of a growing nucleic acid strand. They cannot build from nothing, and they cannot add to the 5-prime end.
Because the two strands of a DNA double helix are antiparallel, this one-way street creates a logistical challenge that requires a team of specialized molecular machines:
- Unwinding: DNA helicase is the enzyme responsible for unwinding the DNA double helix at the replication fork.
- Stabilizing: Once unwound, single-strand binding proteins attach to the unwound DNA to prevent the two complementary strands from reforming hydrogen bonds.
- Relieving Tension: Pulling apart a twisted helix creates intense supercoiling ahead of the fork. Topoisomerase creates temporary nicks in the DNA backbone to relieve supercoiling tension ahead of the replication fork.
- Priming: Because DNA polymerase requires an existing primer strand to initiate the synthesis of a new DNA molecule, DNA primase synthesizes a short RNA primer to provide a starting point for DNA polymerase.
Continuous vs. Discontinuous Synthesis
In prokaryotes, which serve as the canonical model for this process in introductory biology, DNA polymerase III performs the bulk of continuous and discontinuous DNA synthesis during replication.
- The Leading Strand: The leading strand in DNA replication is synthesized continuously in the same direction as the advancing replication fork. It only needs one initial RNA primer, and DNA polymerase III smoothly follows the helicase.
- The Lagging Strand: The lagging strand in DNA replication is synthesized discontinuously in the direction opposite to the advancing replication fork. Because the fork is opening "backward" relative to the strict 5'-to-3' synthesis rule, the lagging strand is built in short bursts. These short segments of DNA synthesized on the lagging strand are known as Okazaki fragments.
The Clean-Up Crew and the Ends of Chromosomes
Once the bulk of the DNA is copied, the RNA primers must be removed. In prokaryotes, DNA polymerase I removes RNA primers from the newly synthesized DNA strands and then replaces the removed RNA primers with DNA nucleotides. Finally, DNA ligase catalyzes the formation of phosphodiester bonds to seal nicks between adjacent DNA fragments, stitching the Okazaki fragments into a single, continuous strand.
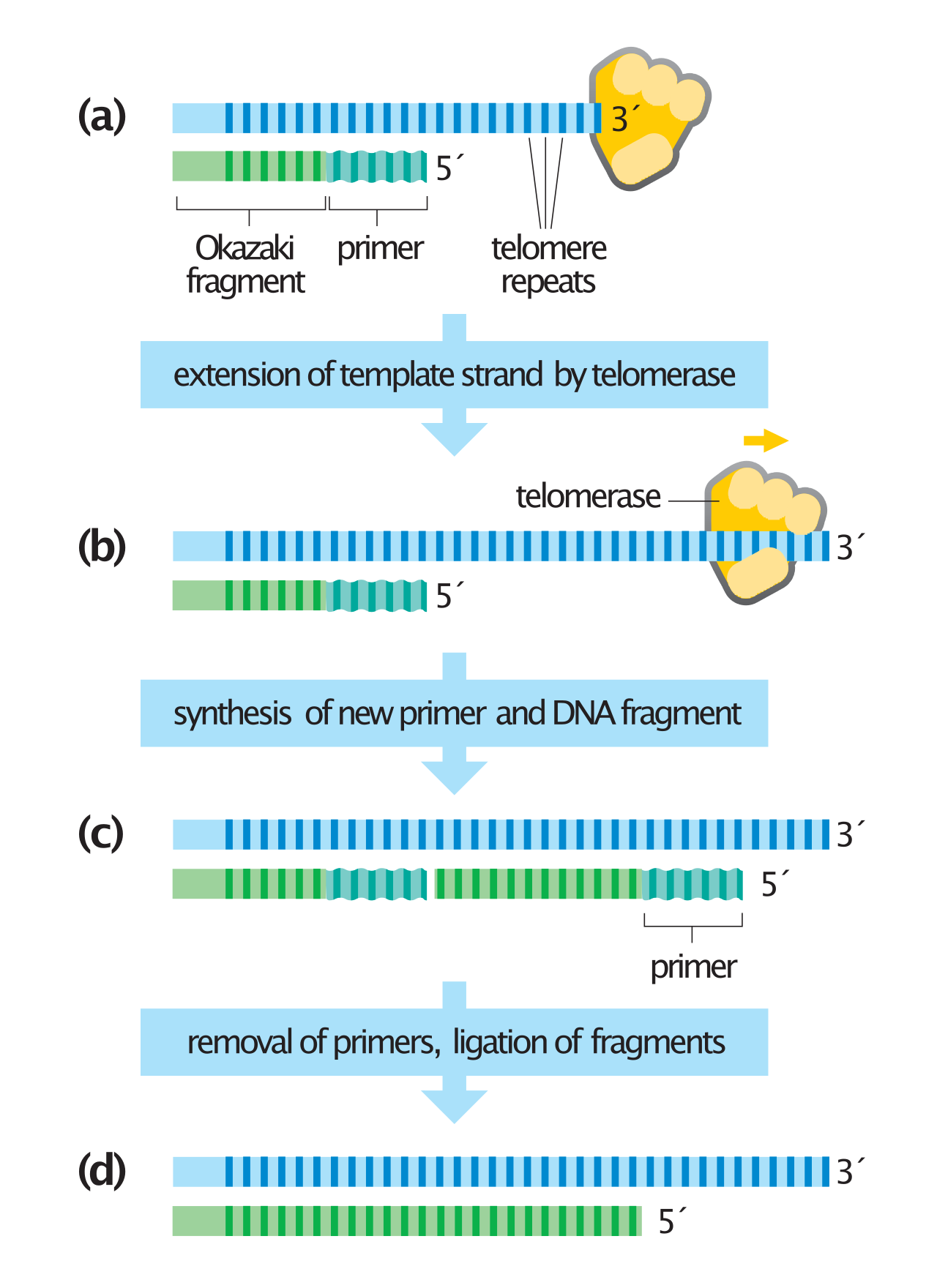
Eukaryotic organisms have linear chromosomes, which introduces the "end-replication problem". When the terminal RNA primer is removed from the lagging strand, there is no upstream 3'-OH group for DNA polymerase to build upon, threatening to shorten the chromosome with every division. Enter telomerase. Telomerase adds repetitive nucleotide sequences to the ends of linear eukaryotic chromosomes. By extending the ends of the chromosomes, telomerase prevents the loss of vital genetic information during repeated cycles of eukaryotic DNA replication.
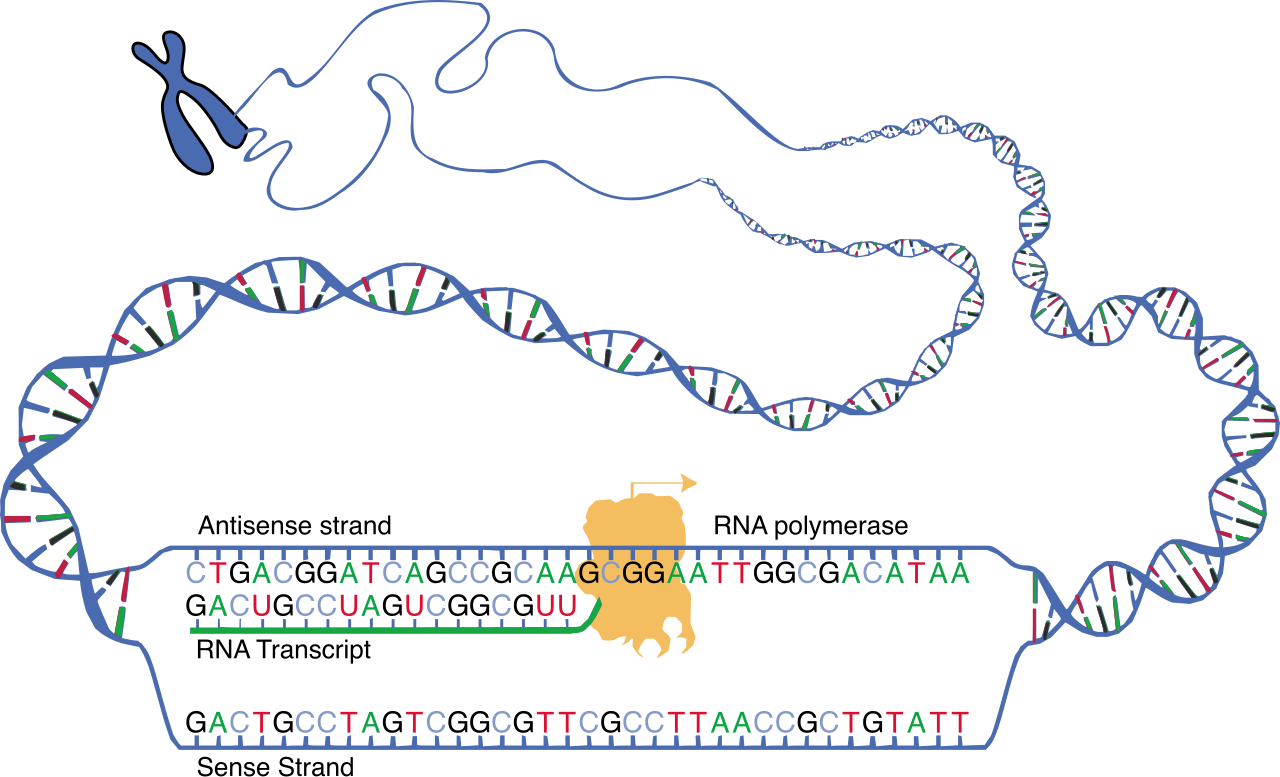
While replication copies the entire library for a new cell, transcription simply checks out a single book. Transcription consists of three primary stages called initiation, elongation, and termination.
Initiation and the Promoter
The star player here is RNA polymerase, which catalyzes the synthesis of a complementary RNA strand from a DNA template. To know where to start, RNA polymerase binds to a specific DNA sequence called a promoter to initiate transcription.
In prokaryotic cells, transcription occurs directly in the cytoplasm, allowing translation to begin even while transcription is finishing. However, eukaryotic cells have a nucleus, adding layers of regulation. In eukaryotic cells, transcription occurs entirely within the nucleus. Furthermore, eukaryotic RNA polymerase cannot bind to the promoter alone. In eukaryotes, general transcription factors must bind to the promoter region before RNA polymerase II can attach. A critical landmark for these factors is the TATA box, a highly conserved DNA sequence found in the promoter region of many eukaryotic genes.
Elongation: Templates and Codes
During elongation, the DNA template strand is read by RNA polymerase in the 3-prime to 5-prime direction. (This allows the new RNA to be built in the 5'-to-3' direction, obeying the universal rule of nucleic acid synthesis).
Exam Tip: Ensure your students understand the nomenclature of the two DNA strands.
- The strand being read is the template strand.
- The non-template DNA strand in transcription is referred to as the coding strand.
Why is it called the coding strand? Because the coding strand shares the exact same nucleotide sequence as the newly synthesized RNA transcript, with one exception: In newly synthesized RNA transcripts, the nitrogenous base uracil replaces the thymine found in the DNA coding strand.
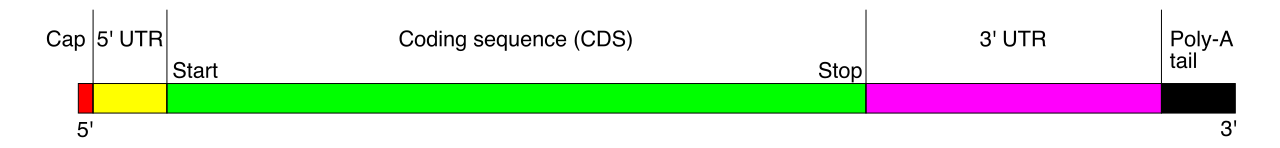
In prokaryotes, the RNA transcript is ready for translation immediately. In eukaryotes, this initial transcript is raw and unpolished. Eukaryotic transcription initially produces a precursor messenger RNA molecule known as pre-mRNA. This eukaryotic pre-mRNA must undergo extensive post-transcriptional processing before exiting the nucleus.
Think of pre-mRNA as rough film footage. Before it hits the theaters (the ribosomes), it needs a title card, a credit sequence, and heavy editing.
1. The 5-Prime Cap
Immediately after transcription begins, a modified 7-methylguanosine nucleotide is added to the 5-prime end of eukaryotic pre-mRNA to form a 5-prime cap. This structure serves two essential purposes:
- The 5-prime cap protects mature mRNA from degradation by exonucleases roaming the cytoplasm.
- The 5-prime cap acts as a recognition signal for ribosome binding during the initiation of translation.
2. The Poly-A Tail
Once transcription is complete, a long sequence of adenine nucleotides is added to the 3-prime end of eukaryotic pre-mRNA to form a poly-A tail. Like a ticking clock, the poly-A tail dictates the lifespan of the mRNA in the cytoplasm, but its immediate function is logistical: the poly-A tail facilitates the export of mature mRNA from the nucleus to the cytoplasm.
3. RNA Splicing
The raw pre-mRNA contains long stretches of genetic "gibberish" that do not code for protein. RNA splicing is the process of excising non-coding nucleotide sequences from a eukaryotic pre-mRNA transcript.
- Introns are the non-coding segments of a pre-mRNA transcript removed during RNA splicing.
- Exons are the coding segments of a pre-mRNA transcript joined together to form the continuous mature mRNA sequence.
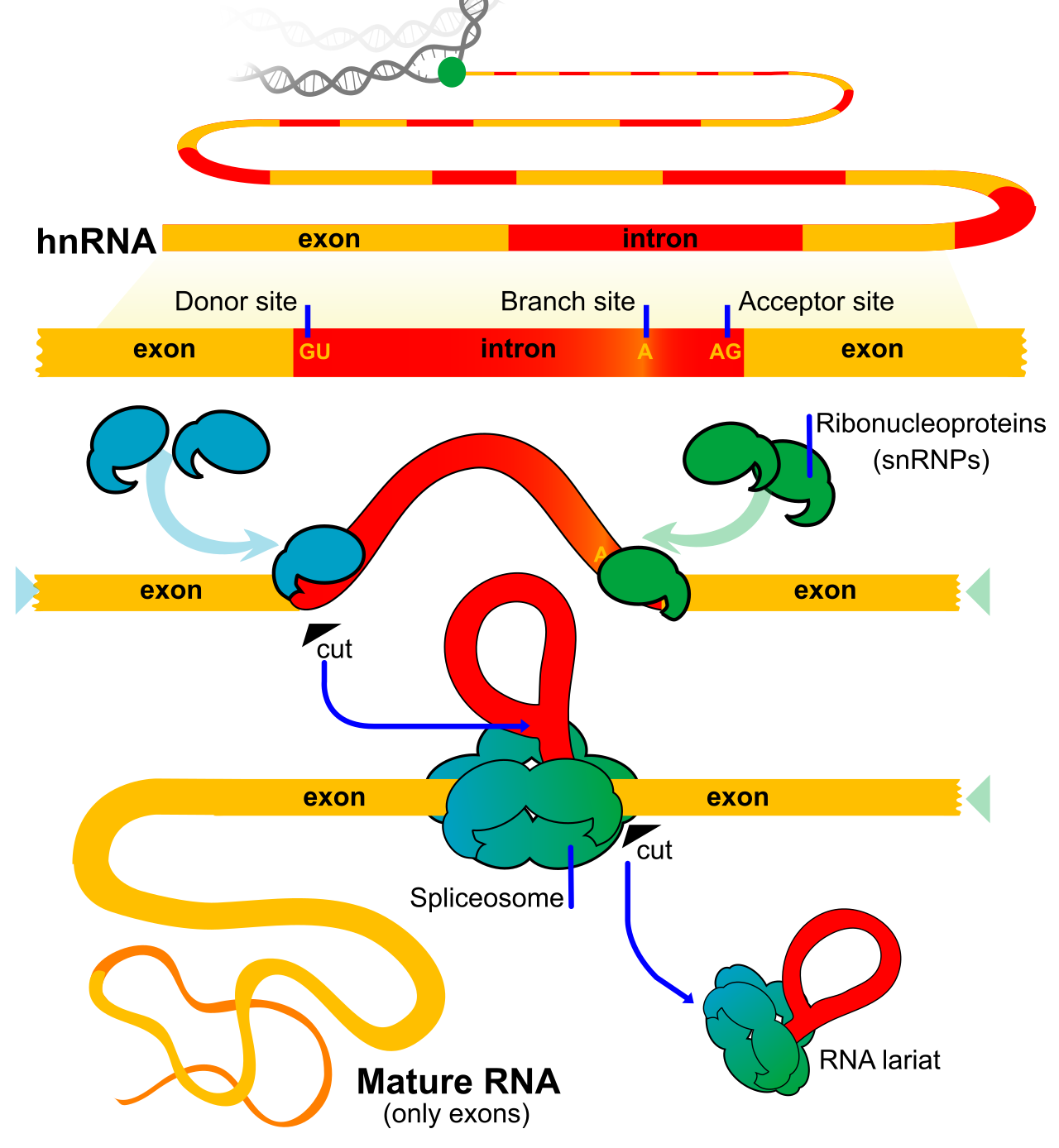
This editing is performed by a massive molecular machine. The spliceosome is a large molecular complex composed of small nuclear ribonucleoproteins (often called snRNPs, or "snurps"). The spliceosome catalyzes the cleavage of introns and the ligation of exons during RNA splicing.
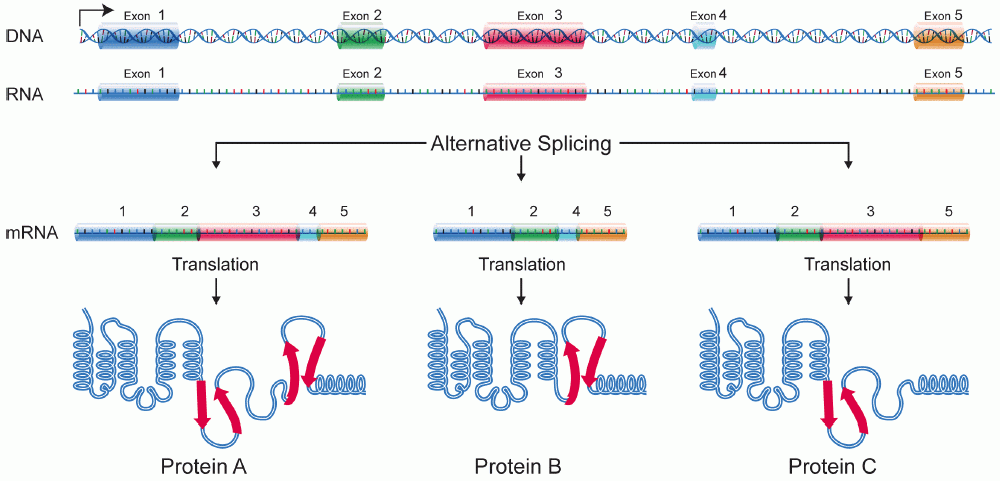
Alternative Splicing: The Engine of Complexity
Why do humans have relatively few genes (around 20,000)—roughly the same number as a microscopic nematode worm—yet exhibit vastly more complexity? The answer lies in how we edit the footage.
Alternative splicing allows a single eukaryotic gene to encode multiple distinct protein isoforms. By treating different segments as introns or exons depending on the cellular context, alternative splicing generates distinct mRNA variants by including different combinations of exons in the final transcript. A gene expressed in a muscle cell might be spliced differently than the exact same gene expressed in a neuron, creating a structural protein in one and a signaling receptor in the other.
Summary Comparison for the Classroom
| Feature | DNA Replication | Transcription |
|---|---|---|
| Primary Enzyme | DNA Polymerase | RNA Polymerase |
| Template | Both DNA strands | Single DNA template strand |
| Primer Required? | Yes (RNA primer) | No |
| Base Pairing | A-T, C-G | A-U, T-A, C-G |
| Product | Double-stranded DNA | Single-stranded RNA |
| Eukaryotic Location | Nucleus | Nucleus |
Mastering the mechanics of replication and transcription gives you, the teacher, the foundational language to explain how a single mutation in a DNA blueprint can ripple through transcription, alter a protein during translation, and ultimately manifest as a dramatic physical trait or genetic disease in the organism.