Translation and Gene Regulation
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Imagine a sprawling, highly efficient manufacturing plant operating autonomously within a microscopic droplet of water. The blueprints are locked securely in a central vault, but the physical construction of products must happen out on the factory floor. To bridge this gap, temporary copies of the instructions are dispatched to specialized molecular machines that read the code and assemble complex physical structures piece by piece. This is not a human engineering marvel, but the fundamental reality of biological life: the translation of genetic information into functional proteins. Understanding how this process is executed and stringently regulated is to understand the physical basis of traits, the origins of metabolic diseases, and the sheer elegance of cellular economy. For an aspiring biology educator, mastering these mechanics is essential for demystifying the molecular world for students who are just beginning to see life through the lens of chemistry and physics.
To build a protein, the cell relies on a cast of specialized RNA molecules, each performing a highly specific role. The process begins with messenger RNA (mRNA), which carries the genetic instructions from the DNA out to the ribosome. If DNA is the master blueprint in the nucleus, mRNA is the disposable photocopy taken to the construction site.
But a sequence of nucleic acids cannot spontaneously assemble a protein. The cell needs delivery trucks. Transfer RNA (tRNA) molecules act as these couriers, engineered to transport specific amino acids to the ribosome.
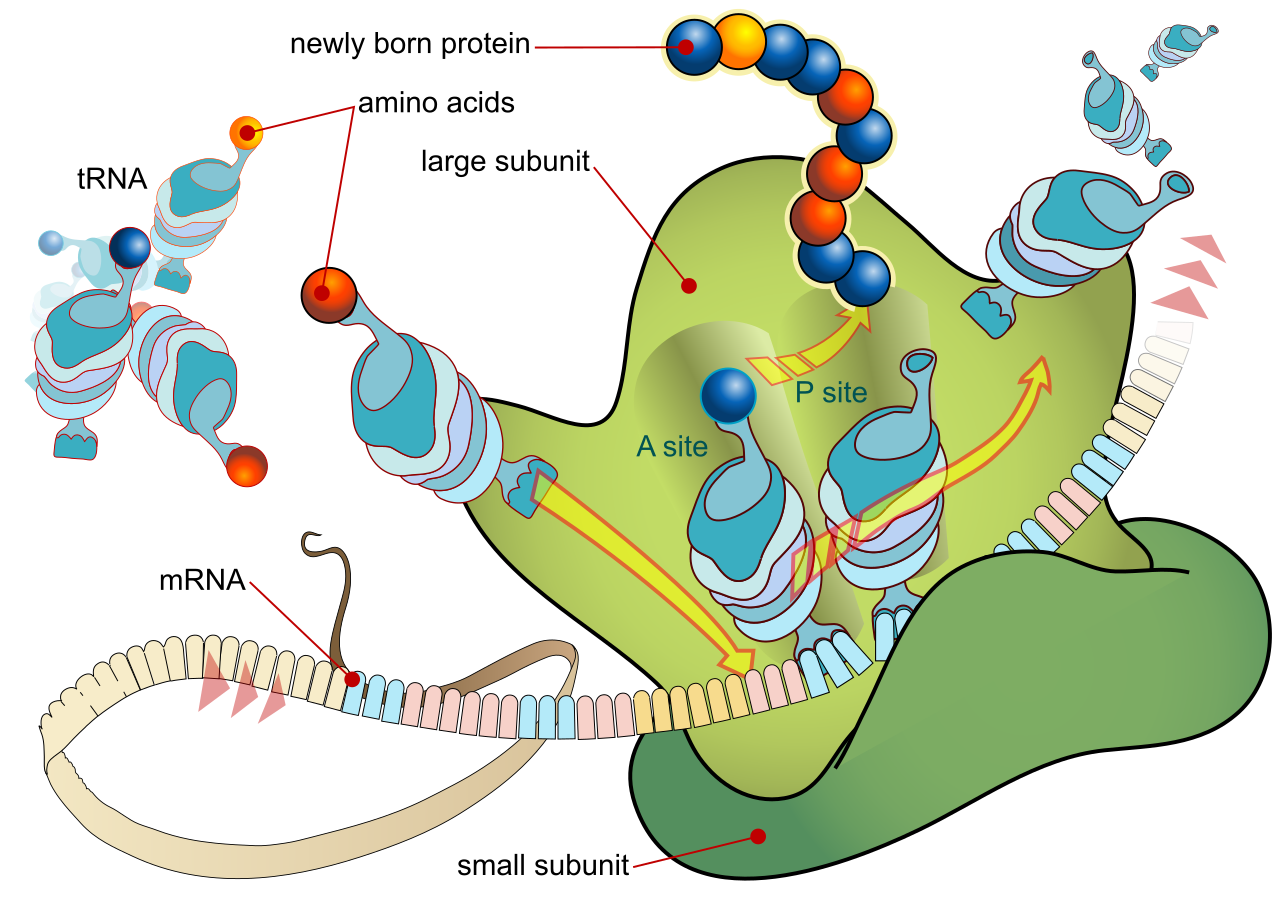
At the center of it all is the ribosome itself, a remarkable molecular machine. A functional ribosome contains a large subunit and a small subunit, which only join together when a protein is about to be made. The actual construction work inside the ribosome—the catalysis of peptide bonds between amino acids—is not performed by a protein enzyme, but by ribosomal RNA (rRNA). It is a striking reminder to your students that RNA is not just a passive information carrier; it is chemically active.
The Ribosomal Assembly Line
The anatomy of the ribosome determines how translation physically happens. The process kicks off when the small ribosomal subunit binds to the messenger RNA transcript. Once the large subunit attaches, the complete factory is ready. The large ribosomal subunit houses three distinct transfer RNA binding sites, creating an incredibly efficient assembly line:
| Ribosomal Site | Function in Translation |
|---|---|
| A site (Aminoacyl) | The entry portal. The A site of the ribosome accepts incoming aminoacyl-tRNA molecules carrying the next amino acid to be added. |
| P site (Peptidyl) | The anchor. The P site of the ribosome holds the transfer RNA carrying the growing polypeptide chain. |
| E site (Exit) | The departure gate. The E site of the ribosome is where empty transfer RNA molecules exit the ribosome to be recharged. |
Deciphering the Blueprint: The Genetic Code
How does the ribosome know which amino acid belongs where? The instructions on the mRNA are written in three-letter words.
A codon is a sequence of three nucleotides on messenger RNA that specifies an amino acid.
To read this, the tRNA comes equipped with a counterpart: an anticodon, which is a three-nucleotide sequence on transfer RNA complementary to a messenger RNA codon.
In your classroom, you will undoubtedly use a genetic code chart, which maps these three-letter messenger RNA codons to specific amino acids. When teaching students to use this chart, emphasize two profound principles of the code:
- The genetic code is redundant because multiple distinct codons can specify the same amino acid. This is a brilliant evolutionary buffer against mutations.
- The genetic code is unambiguous because a single codon specifies only one amino acid. An
AUGwill never mistakenly code for leucine; it means one thing and one thing only.
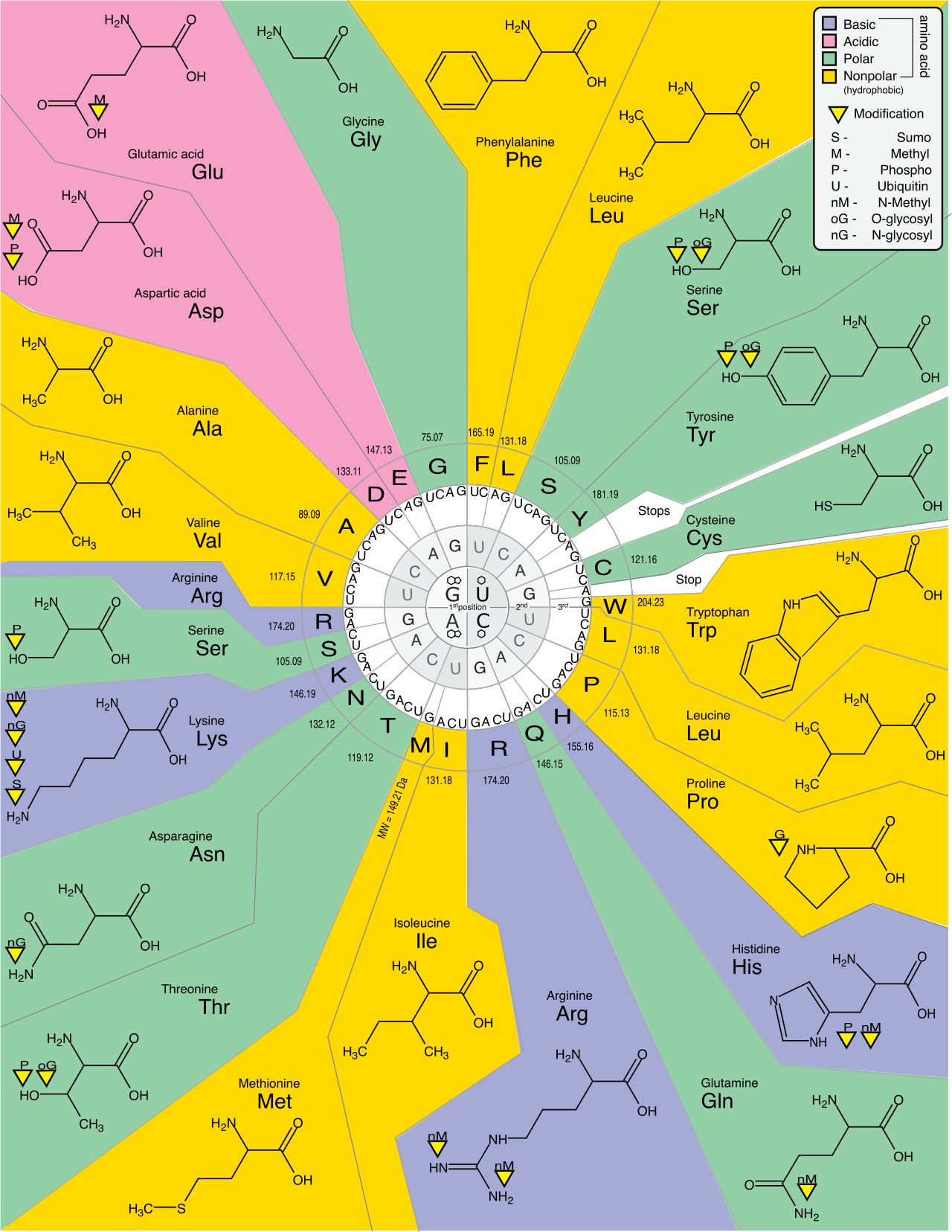
The universal start codon is AUG, which establishes the reading frame and codes for the amino acid methionine. Conversely, there are three stop codons: UAA, UAG, and UGA. Stop codons do not code for any amino acids. Instead, stop codons bind protein release factors to terminate translation, effectively acting as molecular scissors that free the new protein.
The Three Acts of Translation
The chronological process of translation unfolds in three distinct phases:
- Translation initiation involves the assembly of messenger RNA, ribosomal subunits, and an initiator transfer RNA. The reading frame is established by the start codon during translation initiation.
- Translation elongation involves the sequential addition of amino acids to a growing polypeptide chain as tRNAs cycle through the A, P, and E sites.
- Translation termination occurs when a ribosome encounters a stop codon on the messenger RNA transcript.
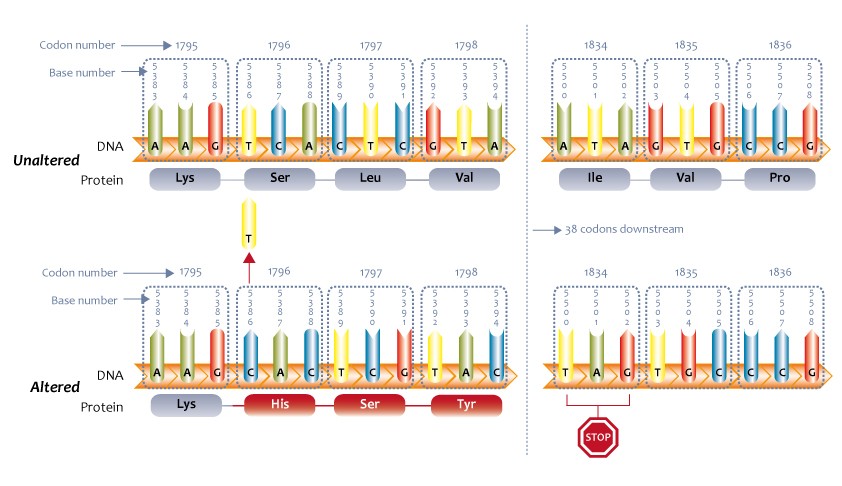
Because translation depends on parsing three-letter blocks, the physical grouping of those letters—the reading frame—is critical. If a mutation inserts or deletes a single nucleotide, it triggers a catastrophic ripple effect. Frameshift mutations alter the reading frame, effectively scrambling the entire message from that point forward. Consequently, frameshift mutations change all amino acid sequences downstream of the mutation site, usually resulting in a completely nonfunctional protein. (A classic classroom analogy: change "THE FAT CAT ATE THE RAT" to "THE FTC ATA TET HER AT" by deleting the first 'A').
Cells do not blindly transcribe and translate every gene they possess. To do so would waste monumental amounts of energy. Bacteria, prioritizing speed and efficiency, handle this masterfully. Prokaryotic gene expression is primarily regulated at the transcriptional level.
Their primary tool is the operon, a cluster of genes under the control of a single promoter in prokaryotes. Think of it as a single power switch controlling every light in a room. Nestled within this setup is an operator, a regulatory DNA sequence within an operon.
The operator is the binding site for repressor proteins. When repressor proteins bind to operator sequences to inhibit transcription, they physically block the track. Repressor proteins physically prevent RNA polymerase from transcribing operon genes.
Let's examine two classic, beautifully opposed systems:
The Inducible Lac Operon: A Logic Gate for Sugar
The lac operon is an inducible operon. By default, it is turned off. The bacteria only want to manufacture the enzymes to digest lactose if lactose is actually present. Thus, the lac operon is activated in the presence of lactose.
When lactose enters the cell, its isomer, allolactose, acts as an inducer by binding to the lac repressor. The binding of allolactose removes the lac repressor from the operator. With the roadblock gone, RNA polymerase races forward, and the bacteria can digest the sugar. Once the sugar is gone, the repressor locks back into place.
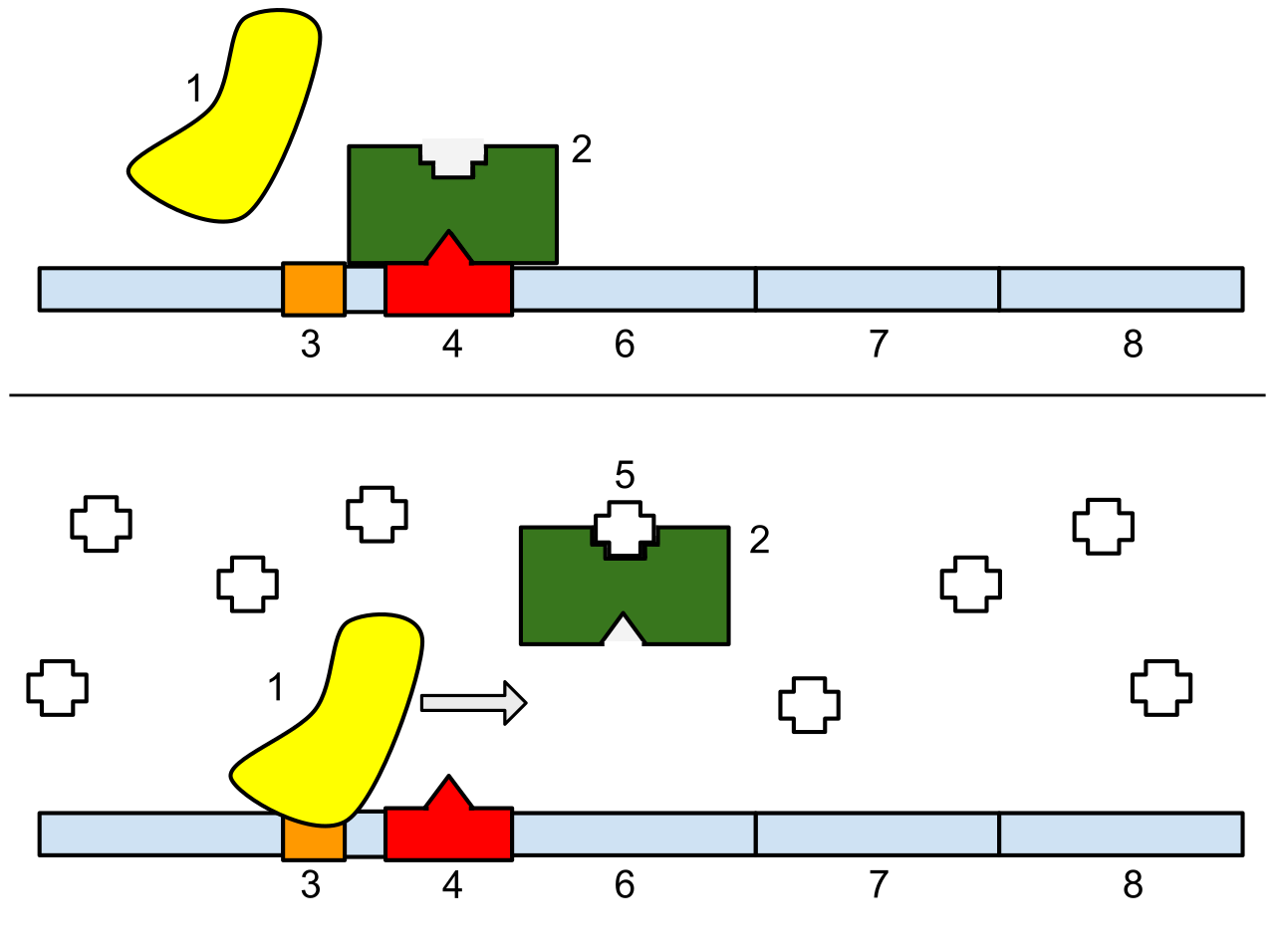
The Repressible Trp Operon: The Molecular Thermostat
Conversely, the trp operon is a repressible operon. It contains the genes to build the amino acid tryptophan. Because the cell constantly needs tryptophan, this operon is normally turned on.
However, if the bacteria absorbs plenty of tryptophan from its environment, it would be wildly inefficient to keep synthesizing it. Therefore, the trp operon is inhibited in the presence of high levels of tryptophan. In this system, tryptophan acts as a corepressor by binding to the trp repressor. Tryptophan binding activates the trp repressor, allowing it to clamp onto the operator and halt production.
While bacteria regulate genes like a simple on/off switch, humans, plants, and fungi must orchestrate the development of trillions of specialized cells, all sharing the exact same DNA. Consequently, eukaryotic gene expression is regulated at epigenetic, transcriptional, post-transcriptional, translational, and post-translational levels.
1. Epigenetic Regulation: Packing the Library
Before RNA polymerase can even attempt to read a gene, the physical structure of the DNA must permit it. Epigenetic regulation involves heritable changes in gene expression without altering the DNA sequence.
Imagine the genome as a vast library. Heterochromatin is tightly packed DNA, effectively locked away in the archives. Heterochromatin is generally inaccessible for RNA transcription. Conversely, euchromatin is loosely packed DNA, sitting out on the reading desks. Euchromatin is readily accessible for RNA transcription.
How does the cell shift between these states? Chemical tags.
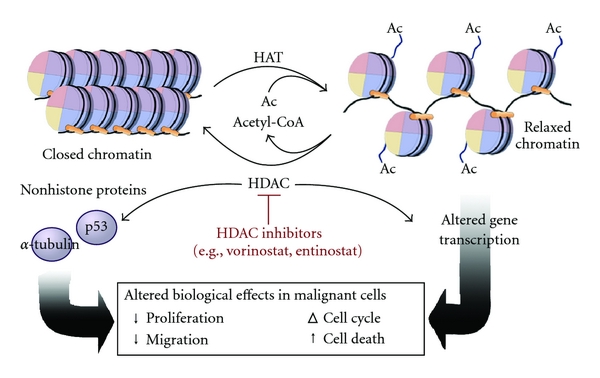
- DNA methylation involves the addition of methyl groups to cytosine bases in DNA. By adding these bulky groups, DNA methylation typically represses gene transcription, packing the DNA tighter.
- Conversely, histone acetylation relaxes chromatin structure. By modifying the histone proteins the DNA wraps around, relaxed chromatin structure promotes gene transcription.
2. Transcriptional Regulation: The Director's Chair
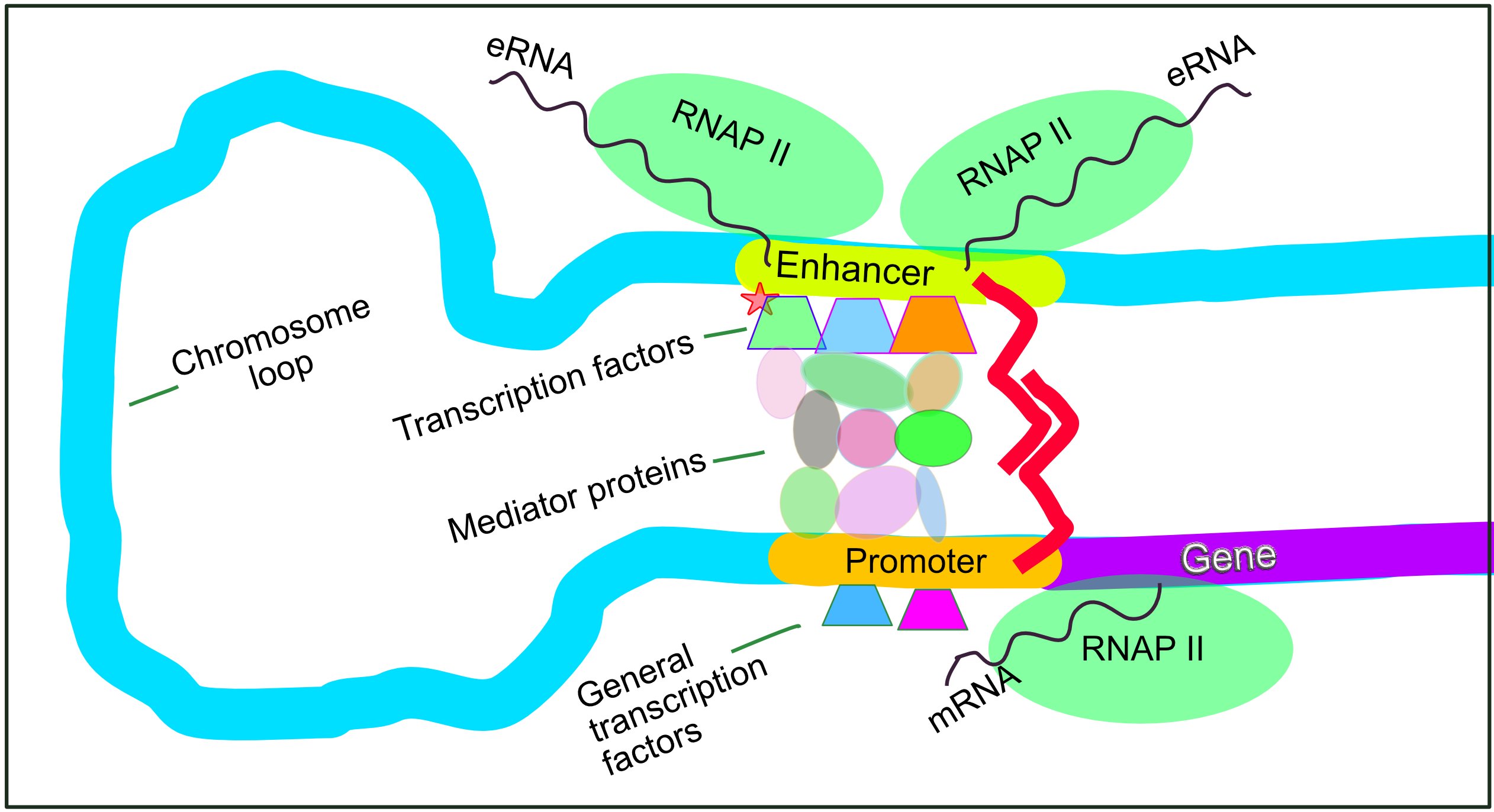
If the DNA is accessible, the next hurdle is initiating transcription. A promoter is a DNA sequence upstream of a gene where RNA polymerase binds. In eukaryotes, the TATA box is a conserved promoter sequence. Unlike in bacteria where RNA polymerase just hops on, eukaryotic regulation requires a VIP pass. General transcription factors must bind to the promoter to recruit RNA polymerase in eukaryotes.
But cells need nuance—a way to express a gene slightly, moderately, or vigorously. Enter enhancers, which are distal regulatory DNA sequences (located far away from the gene itself). Enhancers increase the rate of gene transcription when specific transcription factors called activators bind to enhancer sequences.
To bridge the physical distance between a distal enhancer and the promoter, DNA bending proteins bring enhancer-bound activators close to the promoter region, forming a three-dimensional complex that supercharges transcription. On the opposing end, silencers are regulatory DNA sequences that decrease the rate of gene transcription when specific transcription factors called repressors bind to silencer sequences.
3. Post-Transcriptional Regulation: Editing the Script
Once a eukaryotic gene is transcribed into pre-messenger RNA, the message must be aggressively edited before leaving the nucleus.
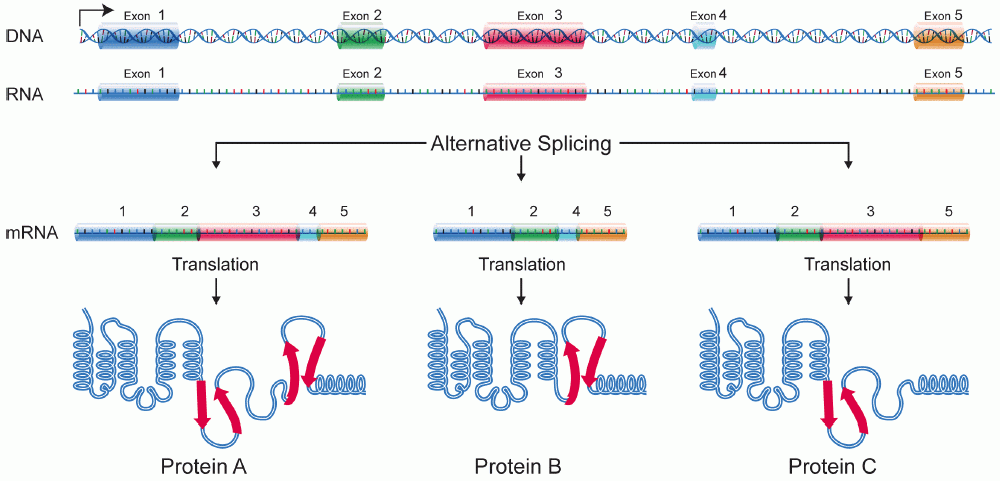
Alternative splicing produces different mature messenger RNA molecules from a single pre-messenger RNA transcript.
This is how humans create hundreds of thousands of different proteins from a mere 20,000 genes. Specialized enzymatic complexes called spliceosomes remove non-coding introns during messenger RNA processing and join coding exons together to form mature messenger RNA. Depending on which exons the spliceosome chooses to include or exclude, the final protein product completely changes.
To survive the treacherous journey through the cytoplasm, the mRNA receives molecular armor: a 5-prime cap is added to protect messenger RNA from degradation, and a 3-prime poly-A tail enhances the stability of messenger RNA in the cytoplasm.
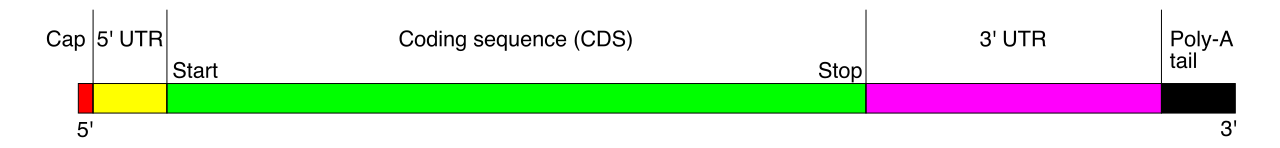
4. Translational Control: The Interceptors
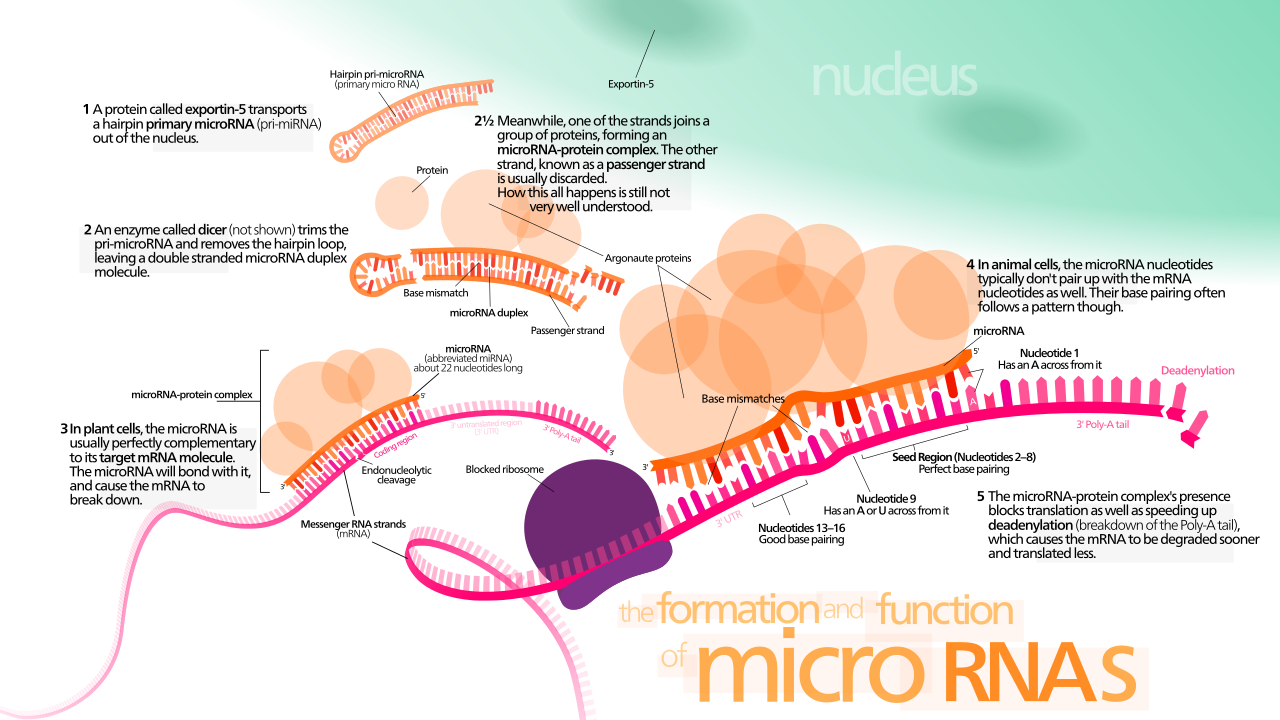
Even with a mature mRNA swimming in the cytoplasm, the cell can still halt translation. MicroRNAs are short non-coding RNA molecules that act as cellular interceptors. Because of their sequence, microRNAs bind to complementary sequences on target messenger RNA transcripts. Once bound, microRNA binding can trigger the degradation of target messenger RNA, or microRNA binding can block the translation of target messenger RNA molecules by getting in the ribosome's way.
5. Post-Translational Regulation: The Final Polish and The Garbage Chute
The polypeptide chain that emerges from the ribosome is rarely a finished, functional protein. Post-translational modifications alter the function of a synthesized protein, or they can alter the stability of a synthesized protein.
For example, protein phosphorylation by kinase enzymes can activate specific target proteins, essentially flipping a chemical switch to turn a resting enzyme into an active one.
Eventually, all proteins become damaged or obsolete. The cell cleans up this hazardous waste using an incredibly targeted mechanism. Ubiquitin molecules are attached to proteins to tag the proteins for destruction. It is the molecular equivalent of drawing a giant red 'X' on a condemned building. Proteasomes are protein complexes that recognize ubiquitin tags. Acting as highly efficient cellular shredders, proteasomes degrade ubiquitin-tagged proteins into short peptides, recycling the amino acids back into the cytoplasm so the beautiful, highly regulated cycle of life can begin all over again.
