Thermodynamics and Metabolism
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
A high-performance internal combustion engine and a marathon runner share a fundamental physical reality: both extract stored potential energy from chemical fuel, convert a fraction of it into kinetic energy to perform mechanical work, and radiate the remainder into their surroundings as waste heat. Thermodynamics is the study of energy transformations in a collection of matter, and it governs every heartbeat, every firing synapse, and every leaf unfurling toward the sun. For a biology educator, framing life not as a static collection of parts, but as a continuous, dynamic flow of energy fundamentally changes how students perceive the natural world. A living cell is not merely a microscopic bag of water and floating proteins; it is a highly active chemical refinery strictly obeying the absolute laws of physics.
To teach cellular energetics effectively, we must first establish the boundaries of the physical universe. Physicists define systems based on how they interact with their environment. An isolated system cannot exchange energy with its external surroundings, nor can an isolated system cannot exchange physical matter with its external surroundings. Think of a perfectly insulated, sealed thermos of hot coffee; it is entirely cut off from the universe outside it.
Living things are vastly different. An open system continuously exchanges energy with its external surroundings, and simultaneously, an open system continuously exchanges physical matter with its external surroundings. We breathe in oxygen, consume food, radiate heat, and exhale carbon dioxide. Therefore, biological organisms operate as open thermodynamic systems.
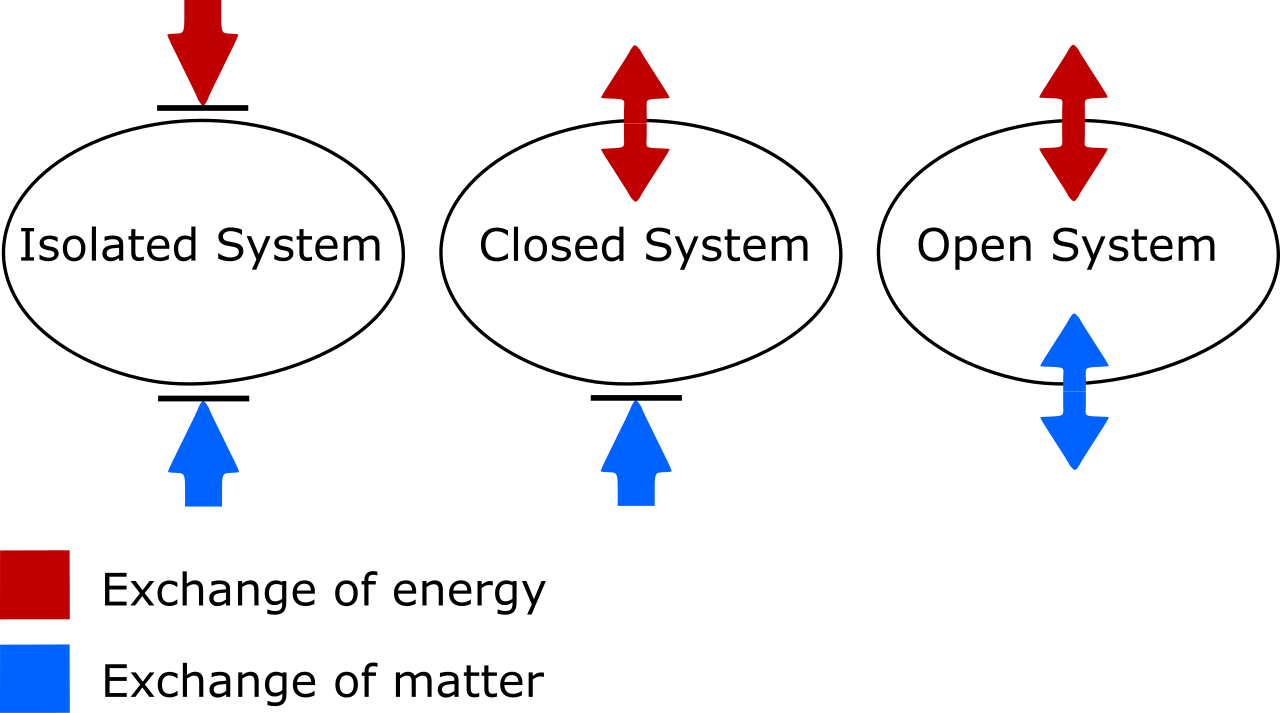
Because organisms are open systems, they are bound by two inescapable universal laws that dictate exactly how energy behaves.
The First Law of Thermodynamics states that the total energy of the universe is constant. Energy cannot be created from nothing, and energy cannot be completely destroyed. However, energy can be transferred from one physical location to another, and energy can be transformed from one physical state to another (such as chemical potential energy converting to kinetic energy).
This first law tells your students that life is a master of conversion, not creation. But the second law demands a tax on every single conversion.
The Second Law of Thermodynamics states that every energy transfer increases the total entropy of the universe.
Entropy is a thermodynamic measure of molecular disorder or randomness. What does this mean for a living, breathing creature? It means perfection is impossible. During every biological energy transfer, some energy becomes unavailable to do cellular work. This is not a biological flaw; it is a physical mandate. This unavailable energy in biological systems is typically dissipated into the environment as heat.
When a student asks why they sweat and radiate heat during a track meet, the answer lies in the Second Law. As muscle cells contract, living cells inevitably convert highly organized forms of potential energy into heat.

This raises a profound philosophical and biological question: if the universe bends inevitably toward disorder, how do complex, highly structured organisms exist at all? The answer is that a living organism maintains its highly ordered internal state by continually increasing the entropy of its surroundings. We eat highly ordered food (like starch) and break it down into scattered, disordered molecules of carbon dioxide and water, dumping heat into the environment. We buy our internal order at the expense of universal chaos.
To determine whether a specific biological reaction will happen on its own, we need a mathematical tool that accounts for both the total energy of the molecules and the entropy tax. Gibbs free energy represents the portion of a biological system's energy available to perform work at a constant temperature and pressure.
For your high school and AP Biology students, the fundamental equation of bioenergetics is paramount:
ΔG=ΔH−TΔS
The change in Gibbs free energy (ΔG) is mathematically defined by subtracting the product of temperature (T) and entropy change (ΔS) from the change in total enthalpy (ΔH).
This equation neatly sorts all biological reactions into two overarching categories:
| Reaction Type | Spontaneity | Free Energy (ΔG) | Biological Role |
|---|---|---|---|
| Exergonic | Exergonic chemical reactions occur spontaneously without an input of external energy. | Exergonic chemical reactions proceed with a net release of Gibbs free energy (ΔG<0). | Breakdown of food; releasing energy to power the cell. |
| Endergonic | Endergonic chemical reactions are non-spontaneous. | Endergonic chemical reactions absorb Gibbs free energy from their surroundings (ΔG>0). | Synthesizing proteins, copying DNA, pumping ions. |
Because cells have a massive checklist of endergonic chores to complete, they employ a brilliant strategy: energy coupling is the biological strategy of using an exergonic process to drive a separate endergonic process. If a reaction costs $50 of free energy, the cell will couple it to a reaction that releases $60 of free energy. The net process is spontaneous, and the cell successfully performs its work.
If we zoom out from single reactions, we see the vast, interconnected web of cellular activity. Metabolism encompasses the totality of an organism's chemical reactions.
Unlike a campfire, which burns glucose into CO2 and H2O in one uncontrolled, fiery flash, biological systems are precise. A metabolic pathway consists of a sequential series of chemical reactions that modify a starting molecule into a specific end product. To control this flow, every individual step in a biological metabolic pathway is catalyzed by a specific enzyme.
Metabolism is fundamentally divided into two opposing forces:
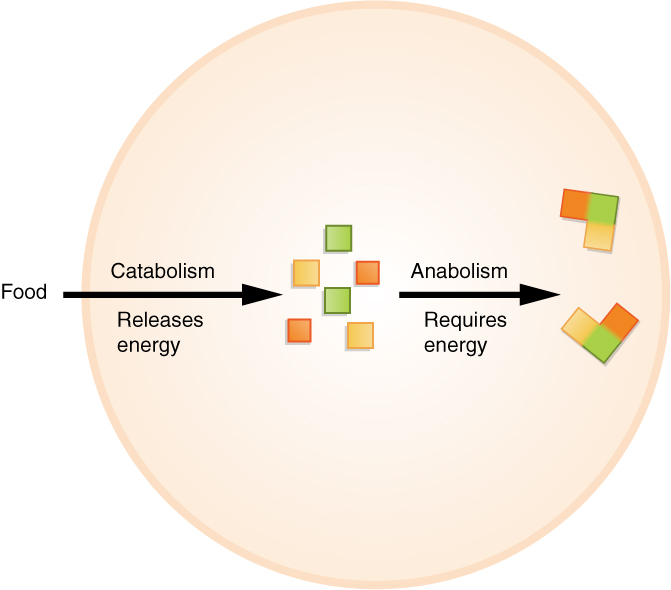
1. Catabolic Pathways (Tearing Down)
Catabolic metabolic pathways release stored energy. They do this because catabolic metabolic pathways break down complex molecules into simpler molecular compounds.
- Classroom Connection: When you teach human digestion or cellular respiration, you are teaching catabolism. Cellular respiration serves as a primary example of a catabolic pathway, systematically tearing apart glucose to harvest its stored potential energy.
2. Anabolic Pathways (Building Up)
Conversely, anabolic metabolic pathways consume energy. The goal here is construction; anabolic metabolic pathways build complex molecules from simpler precursor compounds. For this reason, anabolic pathways are frequently referred to as biosynthetic pathways.
- Classroom Connection: The cellular synthesis of proteins from individual amino acids acts as an anabolic pathway. Muscle growth, DNA replication, and bone development all rely on these energy-intensive pathways.
How exactly is energy harvested from a molecule of glucose and transferred to an anabolic pathway? The secret lies in the movement of electrons.
Oxidation-reduction reactions involve the transfer of one or more electrons from one reactant to another. To keep this straight in the classroom, remind students that electrons carry a negative charge, which is the key to the somewhat counterintuitive terminology:
- Oxidation is defined as the complete or partial loss of electrons from a chemical substance.
- Reduction is defined as the complete or partial addition of electrons to a chemical substance. (Its positive charge is reduced by the added negative electron).
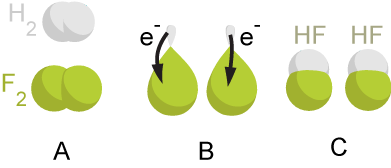
In this energetic dance, roles are highly specific:
- The specific chemical electron donor in a redox reaction is termed the reducing agent (because it causes something else to be reduced).
- The specific chemical electron acceptor in a redox reaction is termed the oxidizing agent (because it snatches the electron, oxidizing the donor).
The Electron Shuttles
Living systems don't just throw electrons across the cytoplasm; they use specialized transport vehicles.
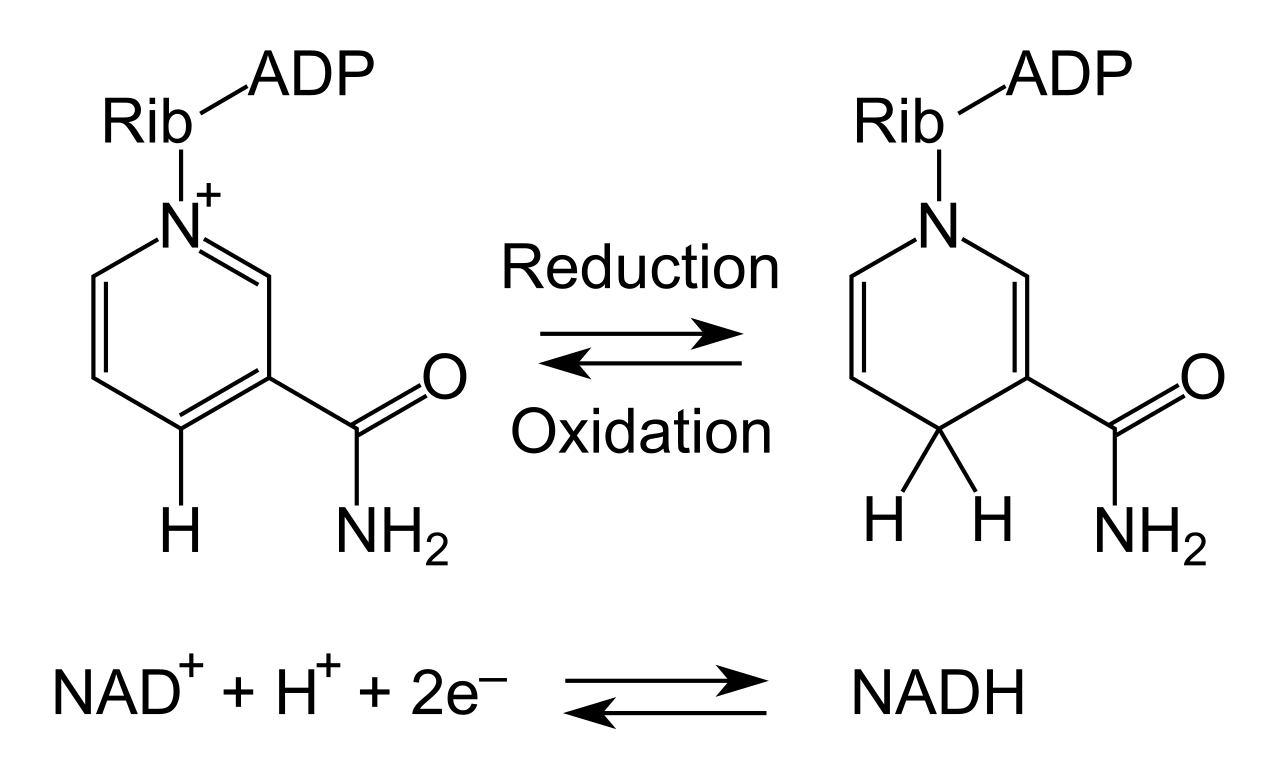
Nicotinamide adenine dinucleotide functions as a highly mobile electron carrier molecule during cellular respiration. It exists in two states depending on whether it is carrying passengers (electrons) or not:
- NAD+ represents the oxidized chemical form of nicotinamide adenine dinucleotide (empty shuttle).
- NADH represents the reduced chemical form of nicotinamide adenine dinucleotide (full shuttle, carrying high-energy electrons).
A secondary, yet crucial, player operates similarly: FAD acts as an oxidizing agent and is reduced to FADH2 during the citric acid cycle.
These molecules are the bridge between the catabolic breakdown of glucose and the ultimate payout of usable cellular energy. They carry their high-energy electron cargo directly to the inner membrane of the mitochondria.
This is where the magic of biological physics truly shines. How do we turn a stream of electrons into physical work?
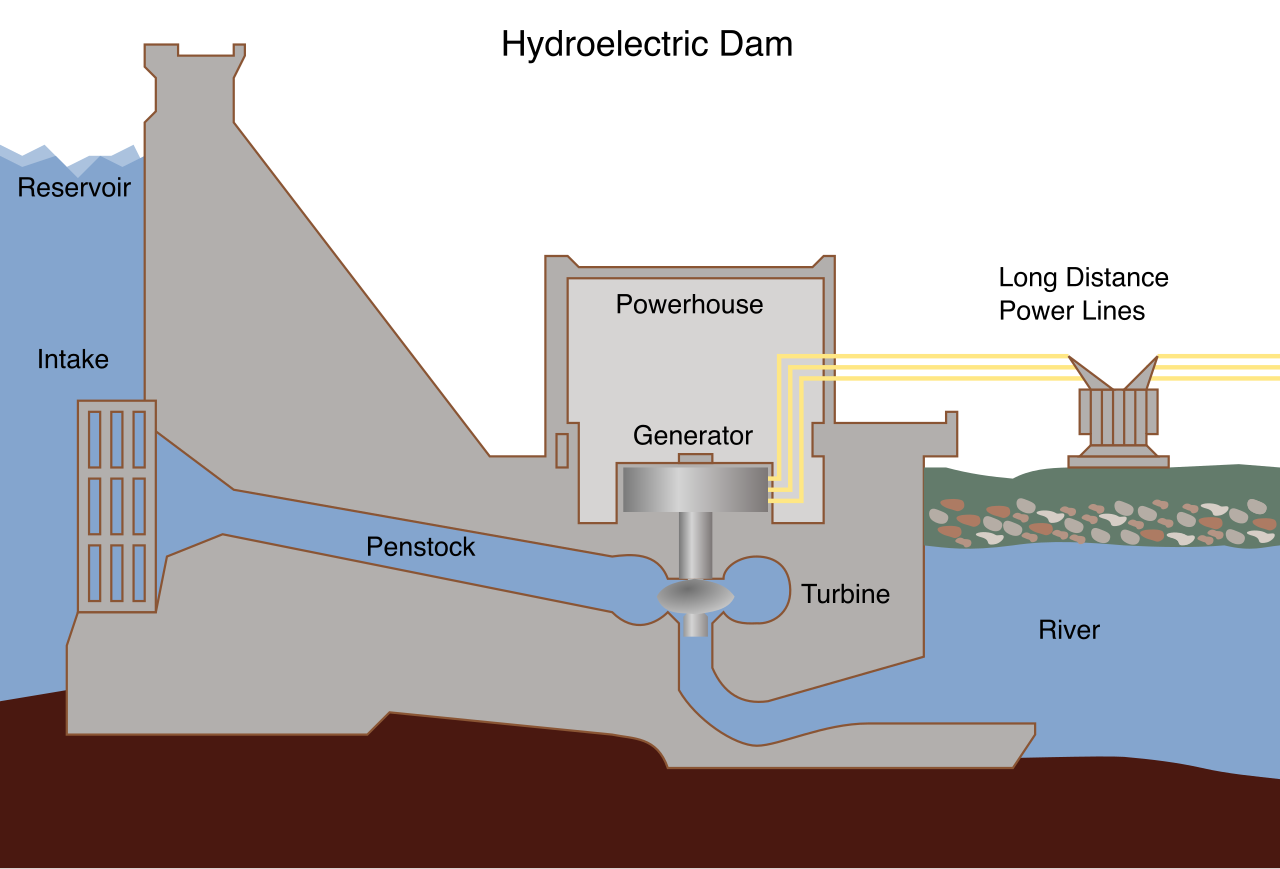
We build a dam.
Chemical concentration gradients established across biological membranes store significant potential energy. Just as water piled high behind a concrete dam desperately "wants" to flow to the lower elevation, molecules packed tightly on one side of a membrane are poised to diffuse across it.
In cellular respiration, this isn't just a chemical gradient; it is an electrochemical one. An electrochemical gradient incorporates a distinct chemical concentration difference across a lipid membrane, but because protons (H+) carry a positive charge, an electrochemical gradient incorporates a measurable electrical charge difference across a lipid membrane.
The Electron Transport Chain
To pump the water behind the dam, the cell uses the high-energy electrons dropped off by NADH and FADH2.
An electron transport chain is a physical series of membrane-bound protein complexes that sequentially transfer electrons via linked redox reactions. As the electrons bounce from one protein complex to the next—moving toward increasingly greedy oxidizing agents until they finally reach Oxygen—they release energy.
Electron transport chains directly couple energetic electron transfers to the active pumping of protons across a membrane. Look closely at this beautiful mechanism: the exergonic fall of electrons is used to do the endergonic work of forcing protons (H+) into the intermembrane space against their will.
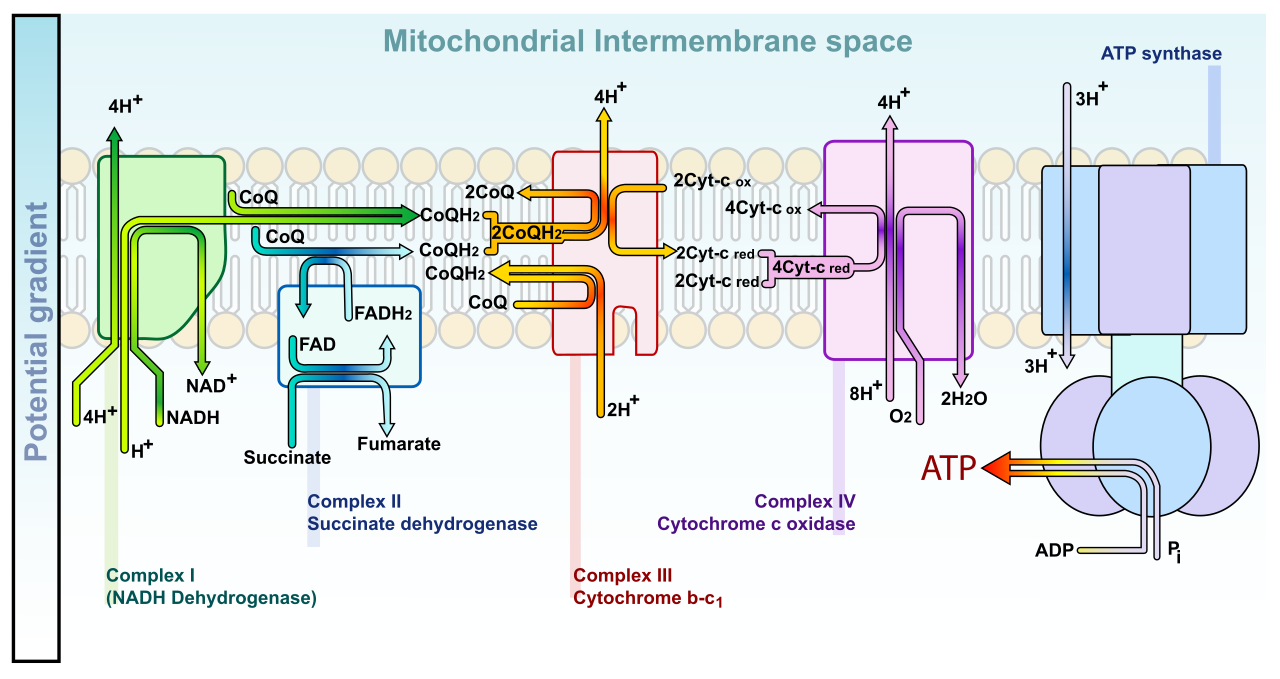
The active pumping of protons across a biological membrane establishes an electrochemical proton gradient. This gradient is volatile and immensely powerful. In fact, proton-motive force describes the capacity of an electrochemical proton gradient to perform cellular work.
Chemiosmosis and the ATP Turbine
Now, the cell lets the dam break—but only through a very specific turbine.
Chemiosmosis is the energy-coupling mechanism utilizing energy stored in a hydrogen ion gradient to drive cellular work. The physical turbine in this scenario is ATP synthase. ATP synthase is a specialized transmembrane enzyme complex.
Much like water rushing through the blades of a hydroelectric turbine, protons rush through the channel in ATP synthase. ATP synthase actively uses the kinetic energy of protons flowing down their electrochemical gradient to synthesize ATP. The physical rotation of the enzyme crushes Adenosine Diphosphate (ADP) and a free inorganic phosphate (Pi) together.
Because it takes energy to force that third phosphate onto the molecule, the cellular synthesis of ATP from ADP and free inorganic phosphate acts as an endergonic reaction.
The Universal Drive Shaft
The result of this incredible, multi-step process is the universal currency of biological energy: ATP.
When the cell needs to flex a muscle fiber, transport an ion, or build a protein, it breaks that high-energy phosphate bond. The chemical hydrolysis of ATP into ADP and free inorganic phosphate acts as an exergonic reaction, releasing the exact burst of free energy required to get the job done.
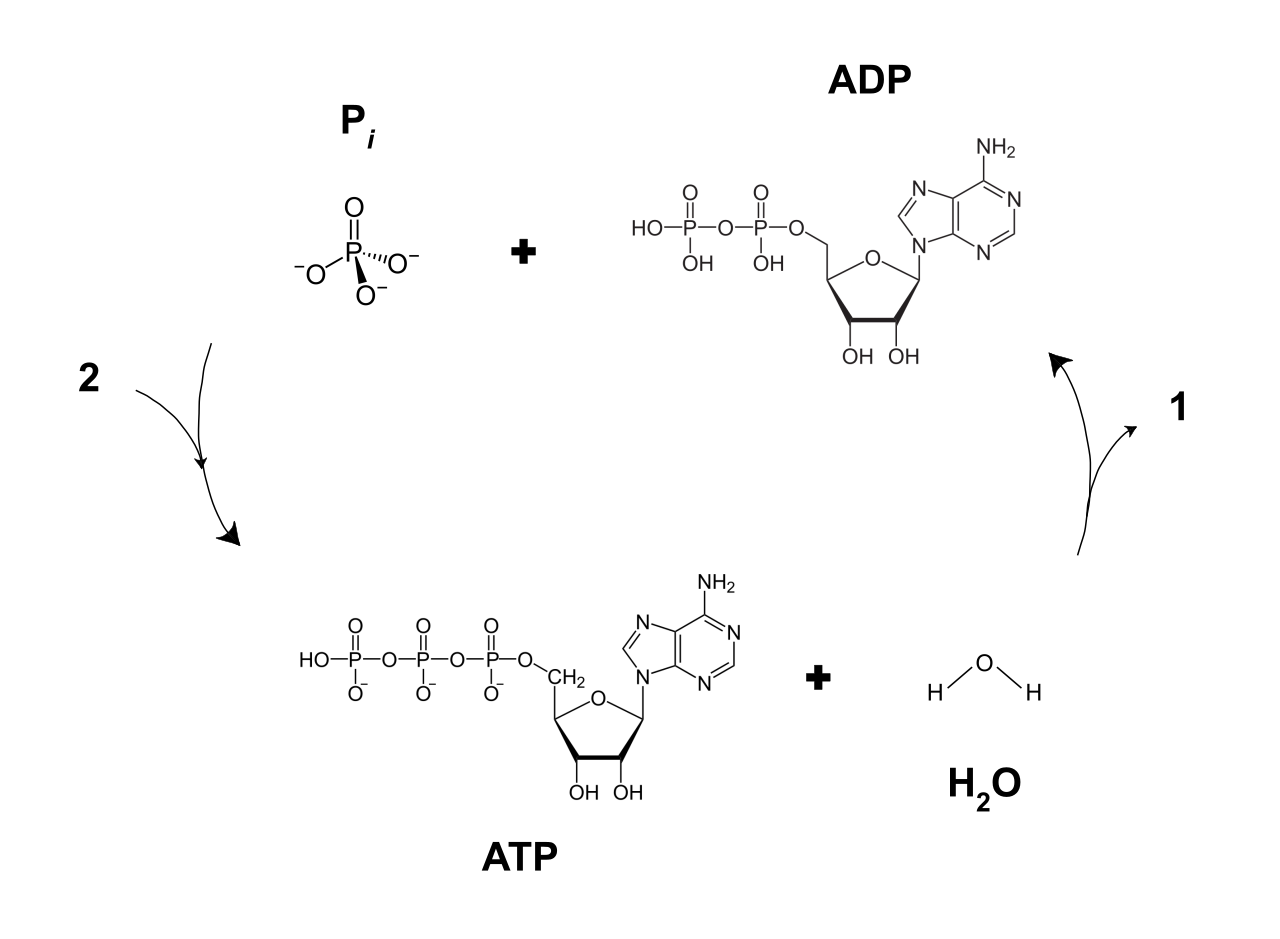
Ultimately, biological systems universally rely on ATP hydrolysis to energetically drive essential endergonic metabolic pathways.
When you stand before your students to diagram cellular respiration or photosynthesis, you are not merely memorizing a biochemical alphabet soup. You are mapping the flow of the universe's energy—from the blinding light of the sun, into the chemical bonds of a leaf, funneled through the gears of oxidation and reduction, temporarily trapped behind a lipid dam, and finally captured in the elegant, vibrating bonds of ATP. This is thermodynamics made flesh.