Plant Characteristics and Structure
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
A towering oak tree or a blade of grass on a suburban lawn is, at its core, a marvel of hydraulic engineering and modular architecture. To survive, a plant must erect a self-assembling, solar-powered skyscraper that pulls massive quantities of water from the soil to its canopy without a single mechanical moving part. It must synthesize its own structural materials from thin air, defend itself against microscopic and macroscopic invaders while rooted firmly in place, and perfectly time its reproductive cycle to the rhythms of the sun and seasons. For a future biology educator, shifting students' perspectives from seeing plants as static background scenery to viewing them as dynamic, fiercely competitive organisms is paramount. To teach botany is to teach the mechanics of survival under profound constraints. By mastering the evolutionary milestones, tissue specializations, and structural anatomy of plants, you equip yourself to explain not just what a plant is, but exactly how it achieves the seemingly impossible.
To understand plant structure, we must first look at the evolutionary hurdles plants had to overcome to conquer dry land. The earliest land plants were limited by their inability to move resources across large distances.
Nonvascular plants lack specialized tissues for transporting water and nutrients. Because they cannot pump water vertically, they are restricted to small sizes and damp environments, relying on simple osmosis and diffusion. Nonvascular plants are collectively known as bryophytes, a group that includes mosses, liverworts, and hornworts. If you look closely at a patch of moss, you will realize that bryophytes lack true roots, lack true stems, and lack true leaves. Furthermore, the gametophyte generation is the dominant phase in the life cycle of nonvascular plants—meaning the green, leafy mat you see on a forest rock is made entirely of haploid cells.

The great evolutionary leap occurred with the development of biological plumbing. Vascular plants, collectively known as tracheophytes, contain specialized transport tissues. This innovation flipped the life cycle hierarchy: the sporophyte generation is the dominant phase in the life cycle of vascular plants. With vascular tissue, plants could finally grow tall, outcompeting their neighbors for sunlight.
Within the vascular plants, reproduction evolved from relying on waterborne spores to producing hardy, easily dispersible seeds:
- Gymnosperms are vascular plants that produce seeds, but these gymnosperm seeds are not enclosed within an ovary (the term translates to "naked seed"). Instead, gymnosperms produce reproductive structures called cones. Conifers (like pine and spruce trees) are a classic example of gymnosperms.
- Angiosperms are also vascular plants that produce seeds, but they took a radically different approach to reproduction. Angiosperm seeds are enclosed within an ovary. Furthermore, angiosperms produce flowers as their primary reproductive structures. After fertilization, the ovary of an angiosperm develops into a fruit, a brilliant evolutionary strategy to bribe animals into dispersing seeds.
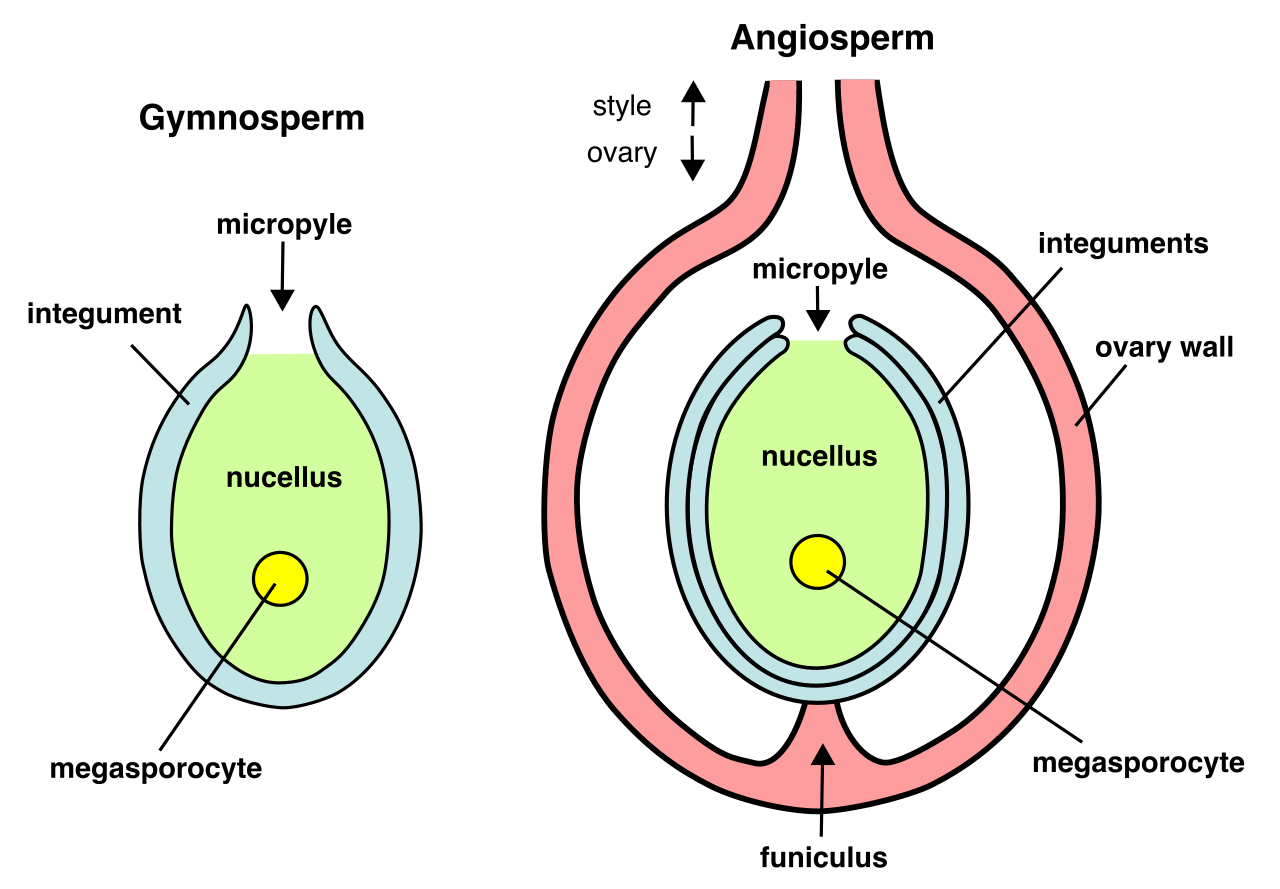
Today, angiosperms dominate the globe and are broadly classified into two main lineages based on their embryonic leaves: monocots and eudicots. We will explore their specific anatomical differences shortly.
From a macroscopic perspective, the plant body is divided into two distinct halves: a root system and a shoot system. The root system functions below ground, while the shoot system handles life above ground. But how does a plant actually grow? Unlike animals, which generally stop growing upon reaching adulthood, plants exhibit indeterminate growth.
This continuous growth is made possible by meristematic tissue.
Meristematic tissue consists of undifferentiated cells capable of continuous cell division. Think of them as the stem cells of the plant world, constantly generating new building blocks.
There are two primary types of meristems:
- Apical meristems are located at the very tips of roots and shoots. Apical meristems facilitate primary plant growth, which results in the lengthening of the plant body. When a tree grows taller or a root reaches deeper, it is the work of apical meristems.
- Lateral meristems facilitate secondary plant growth. Secondary plant growth results in an increase in the girth (or thickness) of the plant body. This is what turns a fragile green sapling into a massive, wide-trunked oak tree.
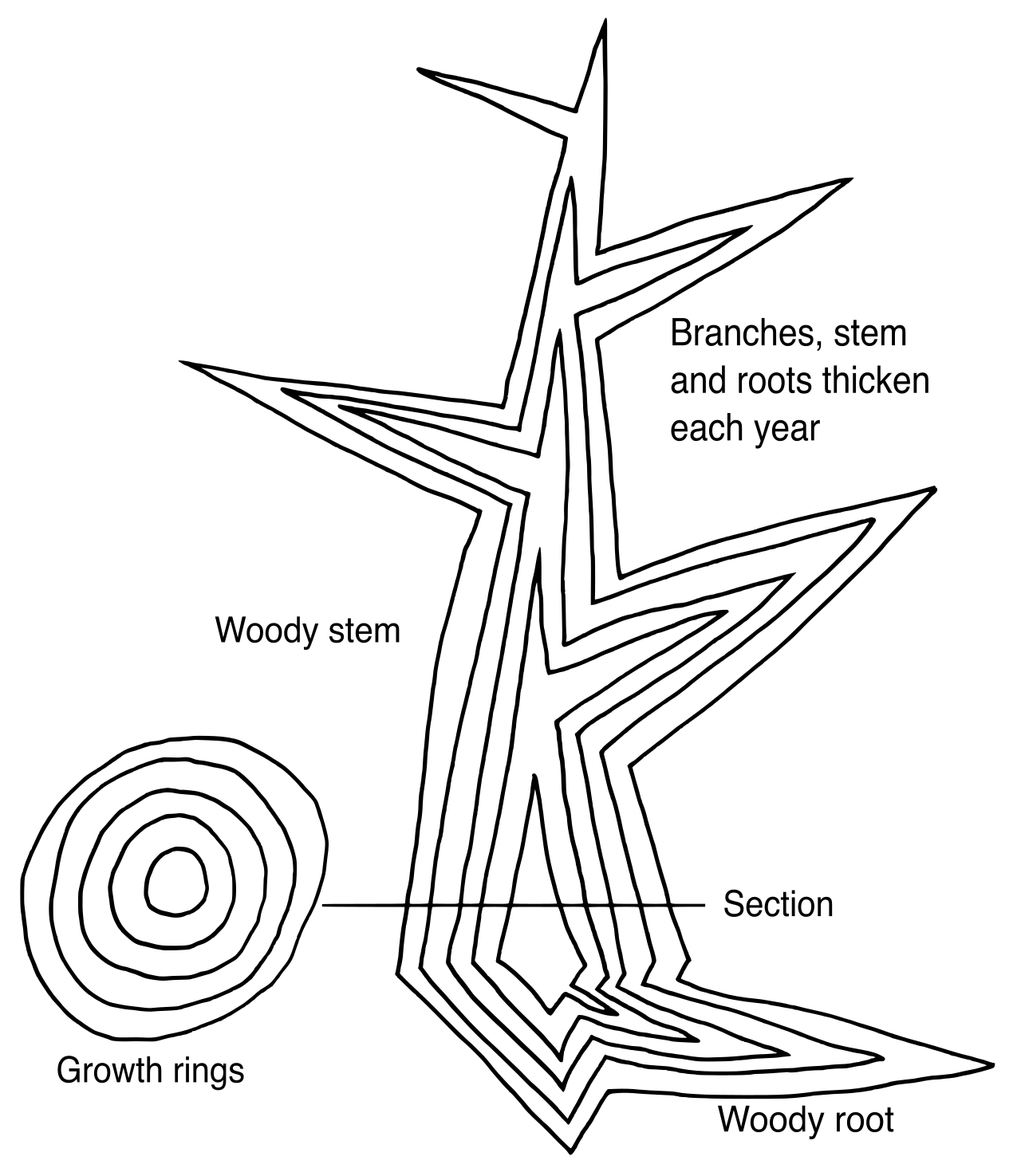
No matter which organ you slice open—a root, a stem, or a leaf—you will find it constructed from three fundamental tissue systems.
1. Dermal Tissue: The Armor
Dermal tissue forms the outer protective covering of the plant. In non-woody plants, the primary dermal tissue is the epidermis. Because plants are in constant danger of drying out, the plant cuticle prevents excessive water loss from surface tissues. The cuticle is a waxy layer secreted by plant epidermal cells, effectively acting as a waterproof sealant.
However, a perfectly sealed plant would suffocate. Therefore, the dermal tissue contains stomata, which are microscopic pores located primarily on leaves. Stomata facilitate gas exchange between the internal plant tissues and the external environment. Because leaving these windows open invites water loss, guard cells flank each stoma to regulate the opening and closing of the pore.
The epidermis also produces specialized structures for interaction with the environment:
- Trichomes are hair-like outgrowths of the plant epidermis. Trichomes can defend plants against herbivores by producing toxic compounds or physical barriers (think of the stinging hairs on a nettle).
- Root hairs are tubular extensions of individual root epidermal cells. Rather than acting as defense, root hairs significantly increase the root surface area for water and mineral absorption underground.
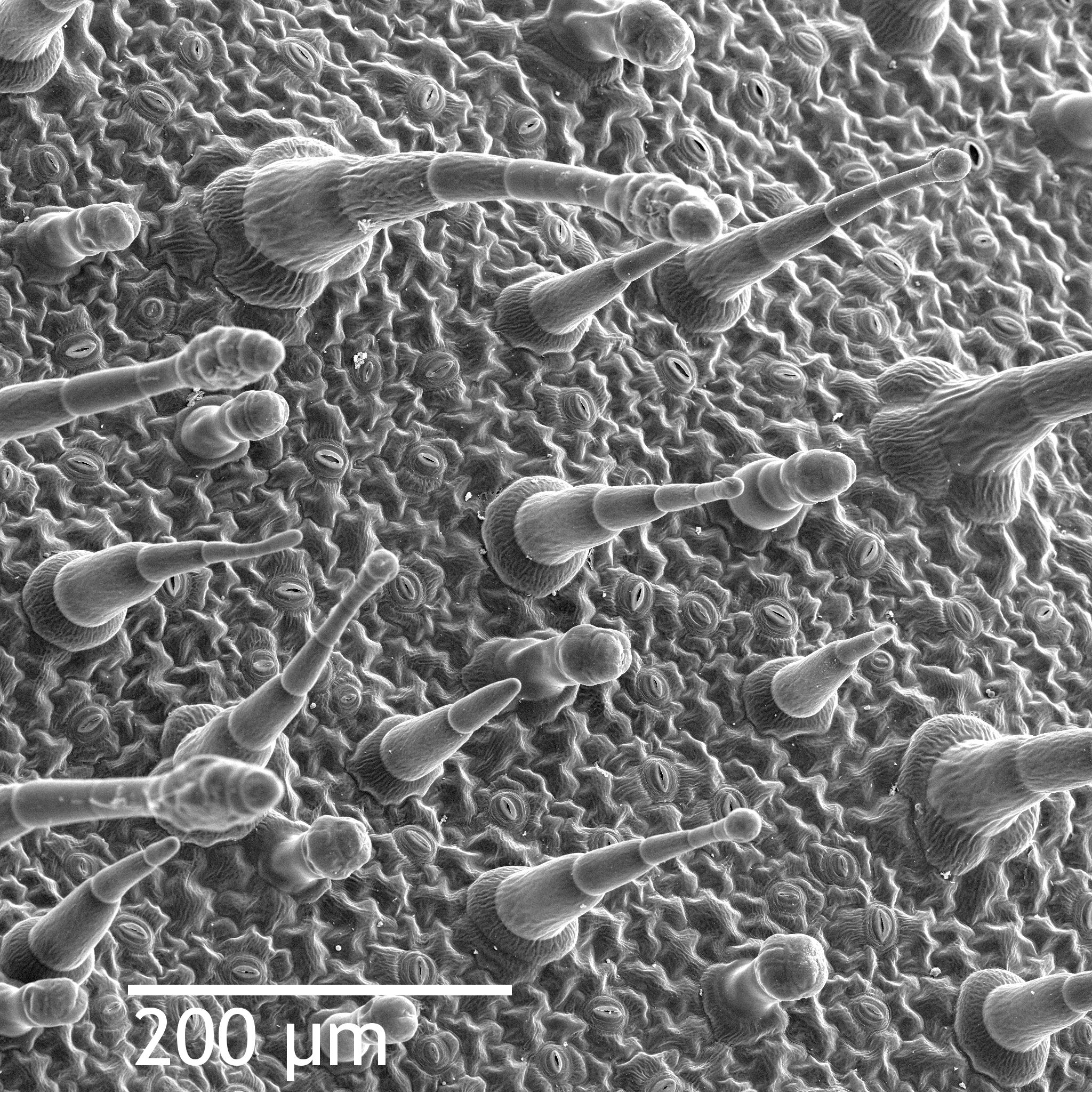
2. Ground Tissue: The Engine and Warehouse
If dermal tissue is the skin, ground tissue is the muscle and fat. Plant ground tissue is responsible for both photosynthesis and carbohydrate storage. It is composed of three distinct cell types:
- Parenchyma cells are the most abundant type of ground tissue in plants. Parenchyma cells have thin primary cell walls and remain alive at functional maturity. They are the versatile "workhorse" cells, carrying out the bulk of metabolic functions like photosynthesis in leaves and starch storage in roots.
- Collenchyma cells provide flexible structural support to growing plant parts. To achieve this flexibility without snapping, collenchyma cells have unevenly thickened primary cell walls and remain alive at functional maturity. When your students ask why celery stalks have those stringy threads that get stuck in their teeth, tell them they are eating collenchyma cells.
- Sclerenchyma cells provide rigid structural support to mature plant parts. Sclerenchyma cells contain thick secondary cell walls fortified with lignin, a highly complex, tough polymer. Because their job is purely structural scaffolding, sclerenchyma cells are dead at functional maturity. When you crack open a tough walnut shell, you are breaking sclerenchyma.
3. Vascular Tissue: The Plumbing
Vascular tissue facilitates the transport of water and nutrients throughout the plant. It operates as a two-way highway system.
Xylem tissue transports water and dissolved minerals from the plant roots to the shoots. The internal architecture of xylem is heavily optimized for fluid dynamics. Xylem tissue contains two types of water-conducting cells: tracheids and vessel elements. Because cytoplasm would obstruct the fast flow of water, tracheids and vessel elements are dead at functional maturity—they function essentially as hollow microscopic pipes.
Phloem tissue, on the other hand, transports synthesized sugars from source tissues (like photosynthesizing leaves) to sink tissues (like developing fruits or roots) throughout the plant. Phloem tissue contains sugar-conducting cells called sieve-tube elements. Because they must actively transport sugars across membranes, sieve-tube elements are alive at functional maturity. However, to leave enough room for sap flow, they lack a nucleus and ribosomes. Consequently, phloem tissue contains supportive cells called companion cells, which are also alive at functional maturity and serve as the life-support system for the sieve-tube elements.
The Root System: Underground Engineering
The plant root system serves three vital functions: it anchors the organism securely in the soil, it absorbs essential water from the soil environment, and it absorbs dissolved minerals from the soil environment.
Root architectures vary by lineage:
- A taproot system consists of one thick primary root with smaller emerging lateral roots. Eudicots typically possess a taproot system (think of a carrot).
- A fibrous root system consists of a dense network of similarly sized roots, which is excellent at preventing soil erosion. Monocots typically possess a fibrous root system (think of common turf grass).

As a root pushes its way through abrasive, rocky earth, it requires protection. The root cap protects the delicate apical meristem as the root pushes through rough soil particles.
Just behind the root cap lies three distinct developmental zones:
- The root zone of cell division, which contains actively dividing meristematic cells.
- The zone of elongation, where plant root cells lengthen to push the root tip deeper into the soil.
- The zone of maturation, where plant root cells ultimately differentiate into specialized tissue types.
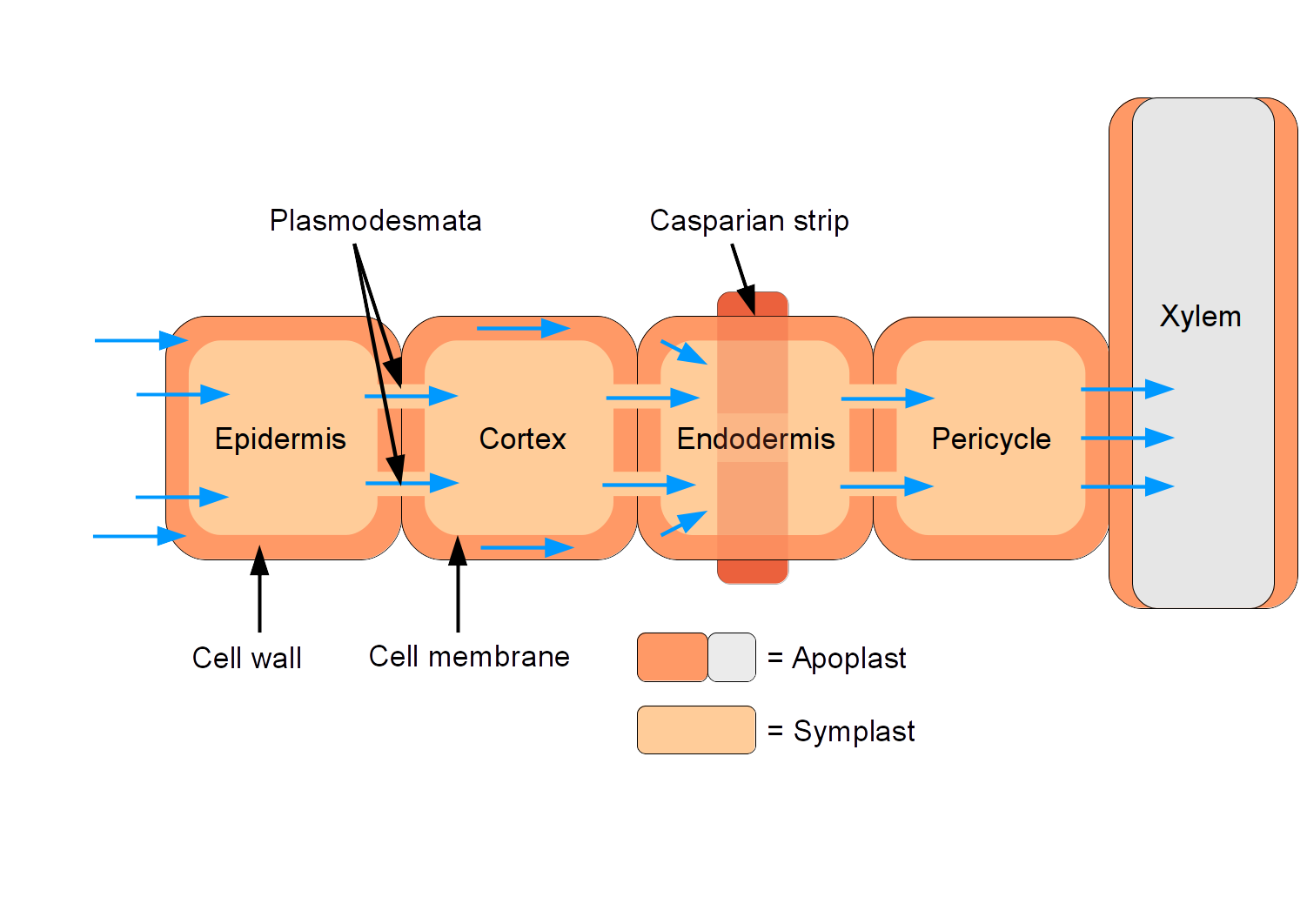
In the zone of maturation, we find a fascinating biological filter. The endodermis is the innermost cellular layer of the root cortex, surrounding the vascular plumbing. Within the endodermis lies the Casparian strip, a water-impermeable waxy band located in the endodermal cell walls of roots.
Why is this important? The Casparian strip acts as a bouncer at a nightclub. It stops water and dissolved substances from slipping passively between cells and forces water to cross a selectively permeable cell membrane before entering the vascular cylinder. This ensures the plant can filter out toxic minerals and regulate its precise internal chemistry.
The Stem: Elevating the Canopy
Plant stems provide physical elevation and support for leaves to maximize light exposure. Stems are built in modular segments. Nodes are specific points on a plant stem where leaves are attached, while internodes are the structural stem segments located between consecutive nodes.
At the intersection of a plant leaf and the stem, you will find an axillary bud. The axillary bud is incredibly important because it has the developmental potential to form a lateral shoot (a new branch).
Internally, stem vascular arrangement acts as a major distinguishing feature between angiosperm clades. In monocot plants, vascular bundles are scattered randomly throughout the stem ground tissue. Conversely, in eudicot plants, vascular bundles are arranged in a distinct peripheral ring within the stem.
The Leaf: The Photosynthetic Factory
Plant leaves serve as the primary anatomical sites of photosynthesis. The macroscopic structure of a typical leaf consists of the leaf blade, which is the broad flat functional part of the leaf designed to catch photons, and the petiole, the stalk structure connecting the leaf blade to the plant stem.
Inside the leaf, sandwiched between the upper and lower epidermis, is the mesophyll, which constitutes the internal photosynthetic ground tissue of a leaf. The mesophyll is highly stratified:
- Palisade mesophyll consists of tightly packed column-shaped cells located right beneath the upper leaf epidermis. Because they are the first line to receive sunlight, palisade mesophyll cells contain the highest concentration of chloroplasts to maximize light absorption.
- Beneath the palisade layer is the spongy mesophyll, which consists of loosely arranged cells separated by extensive air spaces. The air spaces within spongy mesophyll facilitate the rapid diffusion of carbon dioxide and oxygen, allowing the leaf to "breathe" efficiently while photosynthesizing.
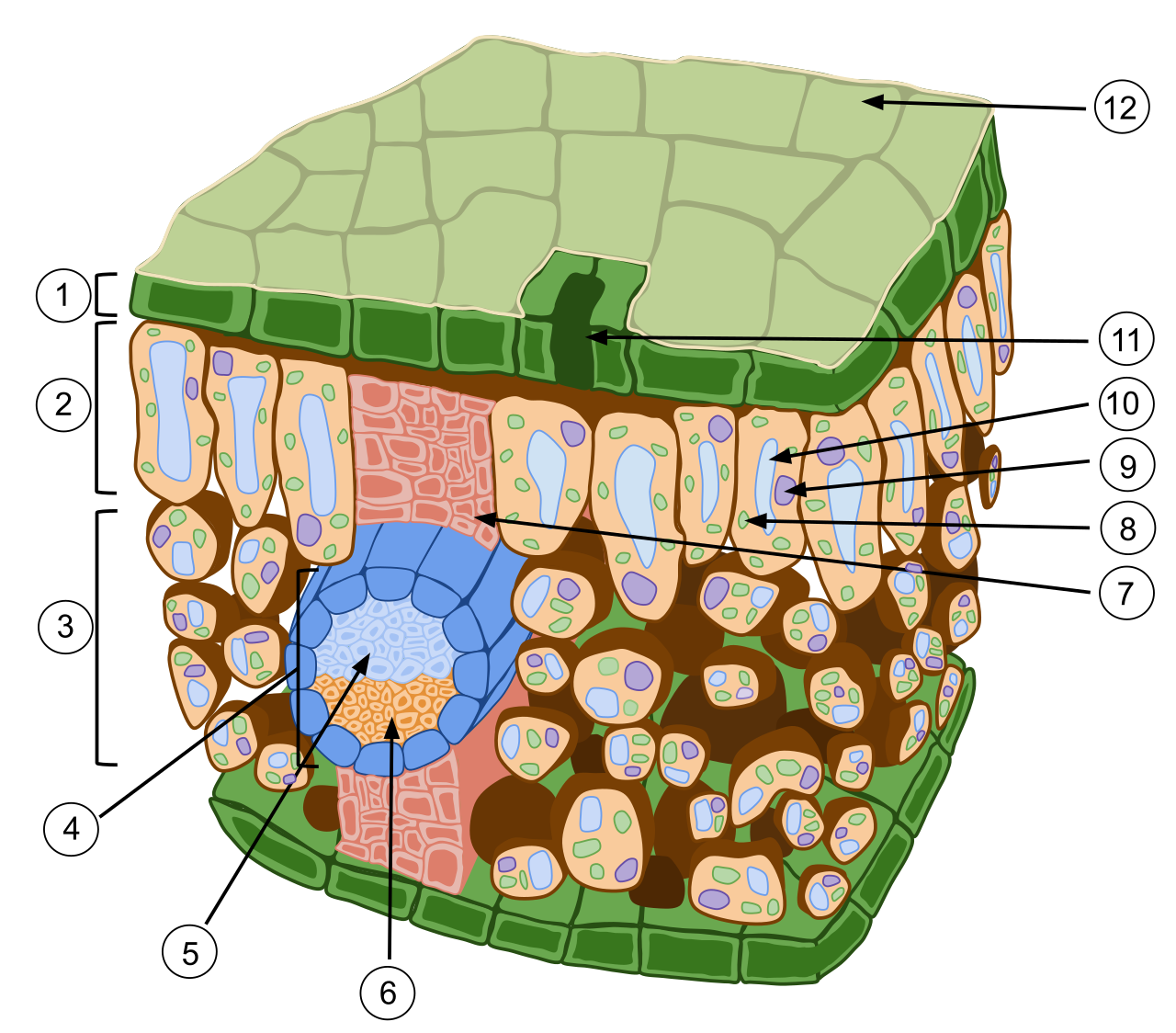
To supply these cells with water and remove their synthesized sugars, leaves contain structural veins. Plant leaf veins are primarily composed of continuous vascular tissue (xylem and phloem) stretching from the stem. These veins also help us classify plants: Monocot plant leaves typically exhibit a parallel venation pattern (like blades of grass or corn leaves), whereas eudicot plant leaves typically exhibit a branched net-like venation pattern (like a maple or oak leaf).
When teaching plant biology, flowers are often the most intuitive entry point for students, but they must be understood scientifically. Flowers are specialized modified shoots containing the angiosperm reproductive organs.
At the very base of the flower is the receptacle, the thickened part of a stem base where all flower organs attach.
The outer structures focus on protection and attraction:
- Sepals are leaf-like structures enclosing and protecting the flower bud before opening. The collective botanical term for all sepals on a single flower is the calyx.
- Just inside the sepals are the petals. Flower petals are frequently brightly colored to visually attract animal pollinators, acting as biological billboards. The collective botanical term for all petals on a single flower is the corolla.
Deep within the corolla sit the reproductive organs.
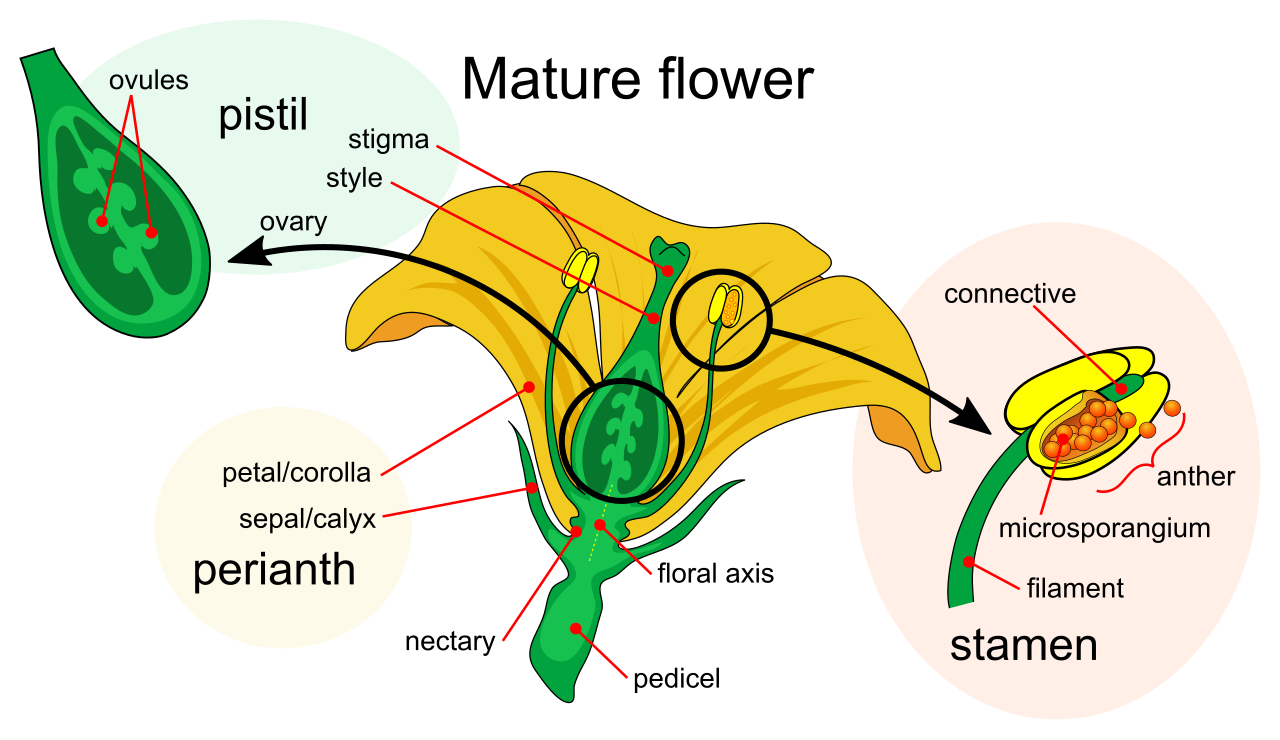
The Male Anatomy: The stamen functions as the male reproductive organ of an angiosperm flower. The stamen includes a supportive stalk called the filament, and it is topped by a terminal sac called the anther. The anther is a microscopic manufacturing plant: the anther produces microspores that ultimately develop into pollen grains. Because plants exhibit an alternation of generations, angiosperm pollen grains actually contain the male gametophytes.
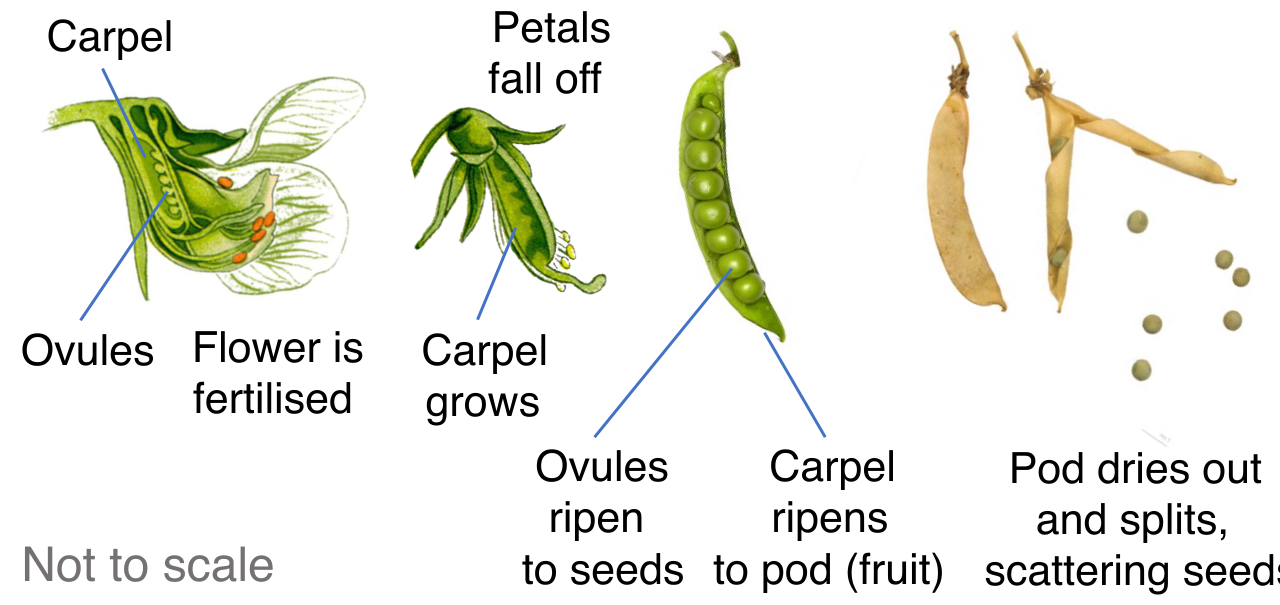
The Female Anatomy: The carpel functions as the female reproductive organ of an angiosperm flower. (Note: A single carpel or a fused group of carpels is botanically termed a pistil).
The carpel is divided into three functional zones:
- The stigma, a sticky apical surface. The stigma serves as the receptive surface for capturing incoming pollen grains.
- The style, a tubular stalk structure through which pollen tubes must grow to reach the reproductive payload.
- The ovary, a swollen basal structure at the bottom of the style. The angiosperm ovary contains one or more ovules.
When a pollen grain successfully fertilizes the female gametophyte inside, the ovule fundamentally transforms. A fertilized plant ovule matures into a seed, safely housing the next sporophyte generation. Concurrently, to ensure that this new seed finds a suitable home far from the shade of its parent, the surrounding ovary of an angiosperm develops into a fruit.
When you prepare your future students for their exams, or simply for a walk through a garden, frame this architecture not as a list of Latin terms, but as a dynamic, interlocking system. From the waxy cuticle to the water-impermeable Casparian strip, and from the dead, hollow tracheids to the vibrant, pollinator-attracting corolla, every millimeter of a plant represents a profound evolutionary triumph over the harsh realities of terrestrial life.