Plant Transport, Reproduction, and Responses
Not sure you’re ready?
Take the ~3-minute readiness diagnostic and see where you stand.
Imagine building a plumbing system capable of defying gravity to lift a column of liquid 300 feet straight up into the sky. Human engineers would require heavy-duty mechanical pumps, immense power grids, and reinforced piping to prevent the system from collapsing under the immense pressure. Yet, the coastal redwood achieves this feat silently, passively, and without a single moving mechanical part. The secret lies in the simple, beautiful physical properties of water and the intricate cellular architecture of plants.
As a future biology educator, you are tasked with revealing the hidden dynamism of the botanical world to your students. To the untrained eye, plants appear as passive scenery. Your goal is to show students that plants are active, highly engineered survival machines engaging in fierce chemical warfare, precise fluid dynamics, and complex reproductive strategies. This guide provides the conceptual foundation you need to master plant transport, reproduction, and environmental responses for your Praxis (5236): Biology: Content Knowledge exam, framed through intuitive analogies that will resonate in your classroom.
Plants manage two opposing traffic flows: water moving up from the roots, and sugars moving down (and around) from the leaves. These processes rely on distinct tissues—the xylem and the phloem—and operate on fundamentally different thermodynamic principles.
1.1 The Pull of the Xylem: Defying Gravity
Water and dissolved minerals are transported upwards from the roots to the shoots through the xylem. Unlike an animal's circulatory system, which relies on a beating heart, xylem transport is primarily a passive process driven by the atmosphere.
This phenomenon is explained by the cohesion-tension theory. To teach this, ask your students to imagine drinking from a straw. When you suck on the straw, you reduce the pressure at the top, and the atmosphere pushes the fluid up. In plants, the "suction" comes from transpiration, which is the loss of water vapor from plant leaves primarily through the stomata.
As water evaporates into the air, transpiration creates a negative pressure that pulls a continuous water column up the xylem. But why doesn't the water column snap under its own weight?
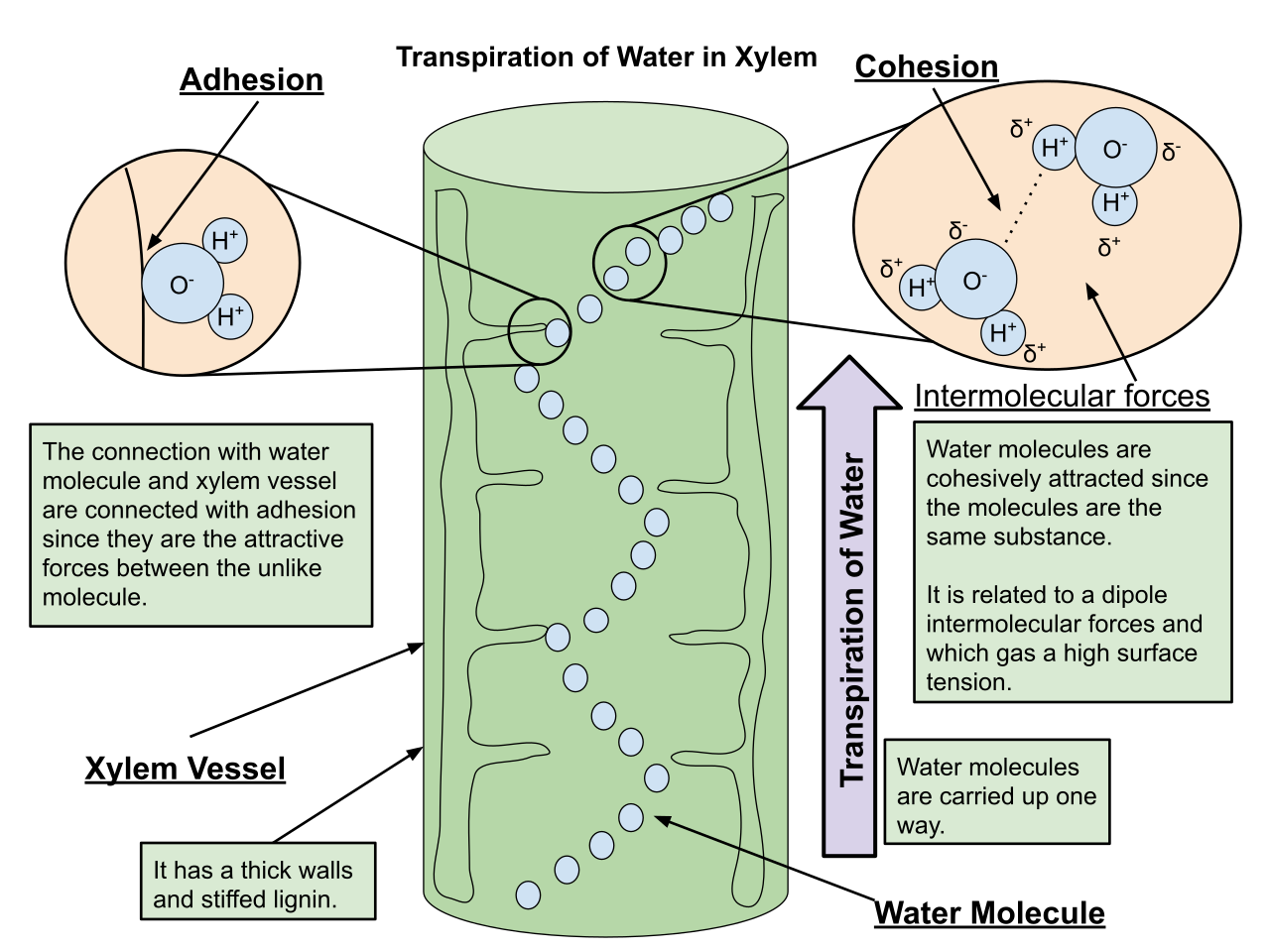
- Cohesion: The attractive force between water molecules (due to hydrogen bonding) helps maintain a continuous water column in the xylem. The molecules "hold hands."
- Adhesion: The attractive force between water molecules and the walls of xylem vessels prevents the water from slipping backward. The molecules "grab the walls."
Classroom Analogy: Think of xylem transport as a chain of monkeys holding hands, dangling from a tree. If you pull the top monkey up, the whole chain moves upward because they refuse to let go of one another (cohesion) and they grip the bark of the tree to stabilize themselves (adhesion).
Water must easily enter and exit the plant's living cells. To facilitate this, aquaporins are channel proteins that facilitate the movement of water across plant cell membranes, vastly increasing the rate of osmosis.
While cohesion-tension does the heavy lifting for tall trees, root pressure can push water up the xylem of short plants. As roots actively pump mineral ions into the xylem, water follows via osmosis, generating a localized hydrostatic pressure that pushes the sap upward.
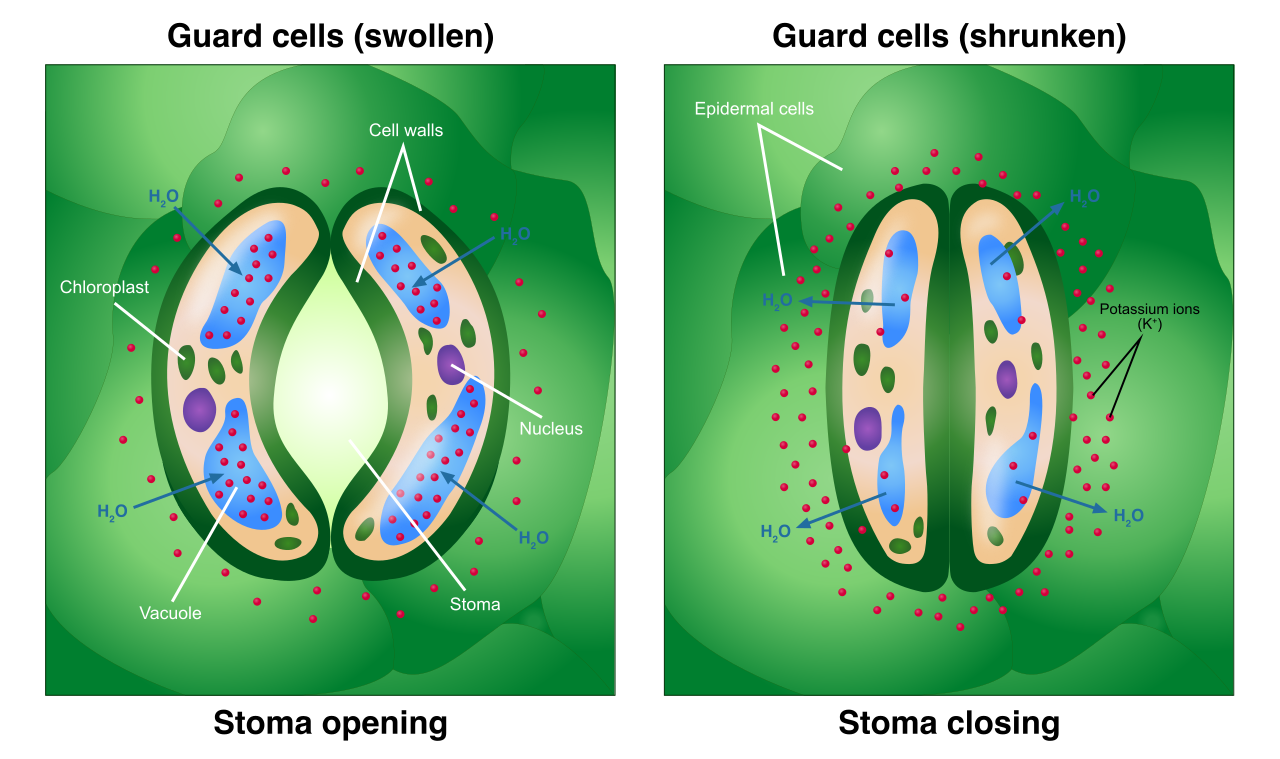
The Regulators: Stomata and Guard Cells
If a plant transpires too much, it desiccates and dies. Thus, stomata are pores on the leaf surface that regulate gas exchange and water loss.
Guard cells flank the stomata and control stomatal opening and closing based on their water volume:
- When guard cells are turgid, stomata open. They bow outward like an over-inflated inner tube, revealing the pore.
- When guard cells are flaccid, stomata close. They lose their bow shape, sealing the pore to conserve water.
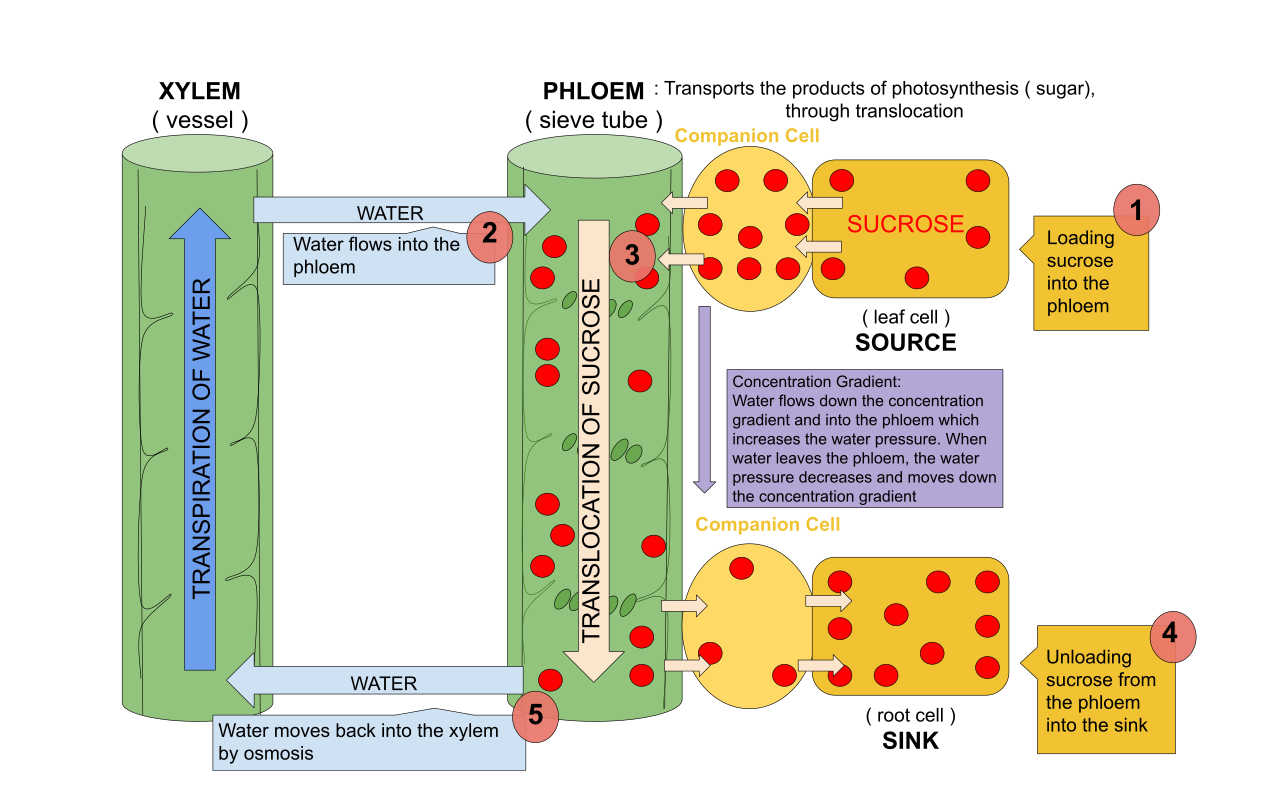
1.2 The Push of the Phloem: Distributing the Wealth
While xylem pulls water up, phloem pushes sap around. Sugars and other organic nutrients are transported throughout the plant via the phloem.
The mechanism governing this is the pressure-flow hypothesis. Instead of relying on evaporation, phloem sap moves from a sugar source to a sugar sink.
- A sugar source is a plant organ that is a net producer of sugar such as mature leaves (via photosynthesis).
- A sugar sink is a plant organ that is a net consumer or storer of sugar such as roots or developing fruits.
This process requires energy. Active transport is required to load sucrose into phloem sieve-tube elements at the sugar source. Once the sucrose is packed into the phloem, the local environment becomes hypertonic. Consequently, this high sucrose concentration in the phloem draws water in from the adjacent xylem via osmosis.
This influx of water is the engine of phloem transport: the increased water volume in the phloem at the source creates positive pressure that drives sap flow towards the sink, much like squeezing a tube of toothpaste forces the paste to the other end.
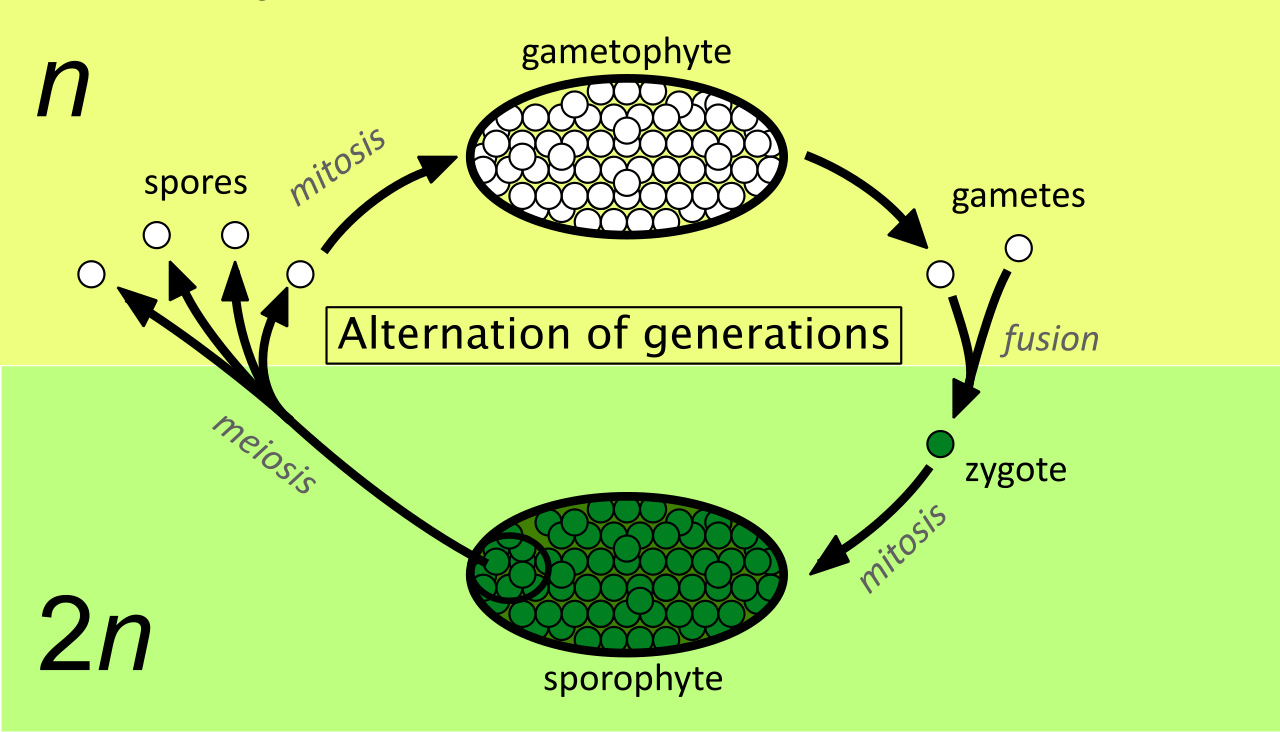
Plant reproduction represents one of the greatest conceptual hurdles for biology students because it fundamentally differs from human reproduction. Plants exhibit a life cycle called alternation of generations, which involves both a multicellular diploid stage and a multicellular haploid stage.
To translate this for students, tell them to imagine a world where human sperm and eggs didn't just fuse immediately, but instead grew into independent, free-living, multicellular organisms that existed solely to eventually produce gametes.
2.1 The Two Generations
- The sporophyte is the multicellular diploid stage in a plant life cycle. By definition, the sporophyte produces haploid spores through the process of meiosis.
- The gametophyte is the multicellular haploid stage in a plant life cycle. Crucially, because it is already haploid, the gametophyte produces haploid gametes through the process of mitosis.
Over evolutionary time, the balance of power between these stages shifted:
- In mosses and other bryophytes, the gametophyte is the dominant photosynthetic generation. (The green, carpet-like moss you see is haploid tissue).
- In ferns, gymnosperms, and angiosperms, the sporophyte is the dominant generation. (The giant oak tree, the rose bush, and the fern frond are all diploid).
2.2 Angiosperm Anatomy and Gametogenesis
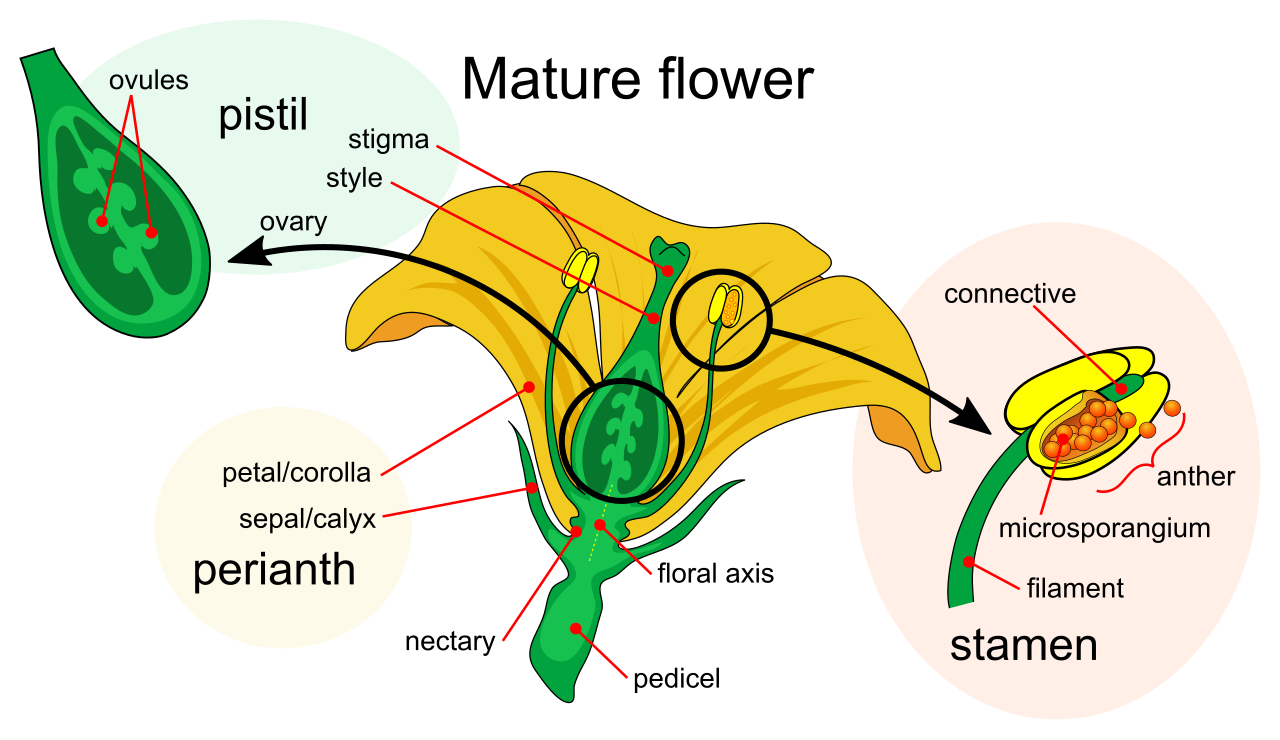
Angiosperms are flowering plants that produce seeds enclosed within a fruit. The flower is a highly modified shoot designed exclusively for sex.
Male Anatomy:
- The anther is the pollen-producing part of a flower stamen.
- Deep inside, microsporangia within the anther produce microspores that develop into male gametophytes.
- Ultimately, a pollen grain represents the male gametophyte in seed plants. It is a tiny, durable, multicellular haploid organism that flies through the air.
Female Anatomy:
- The ovary of a flower contains one or more ovules.
- Inside, megasporangia within the ovule produce megaspores that develop into female gametophytes.
- The embryo sac is the female gametophyte in angiosperms, typically consisting of just seven cells buried deep within the flower.
2.3 Pollination and the "Package Deal" of Fertilization
Pollination is the transfer of pollen from an anther to a receptive stigma. Plants utilize different vectors to achieve this, reflected in their morphology:
- Wind-pollinated plants typically produce large amounts of lightweight pollen and have inconspicuous flowers. (Think of grasses and oak trees, which cause seasonal allergies).
- Animal-pollinated plants often have brightly colored petals, nectar, or distinct scents to attract pollinators.
Once pollen lands on the stigma, a remarkable event occurs: double fertilization is a reproductive mechanism unique to angiosperms. The pollen tube delivers two sperm cells to the embryo sac:
- In double fertilization, one sperm fertilizes the egg to form a diploid zygote.
- In double fertilization, a second sperm fuses with two polar nuclei to form a triploid endosperm.
Why Double Fertilization? It prevents plants from wasting resources. The endosperm provides nourishment to the developing plant embryo, but it only forms if an egg is actually fertilized. It is the ultimate biological "package deal"—one sperm creates the baby, the other creates the baby's packed lunch.
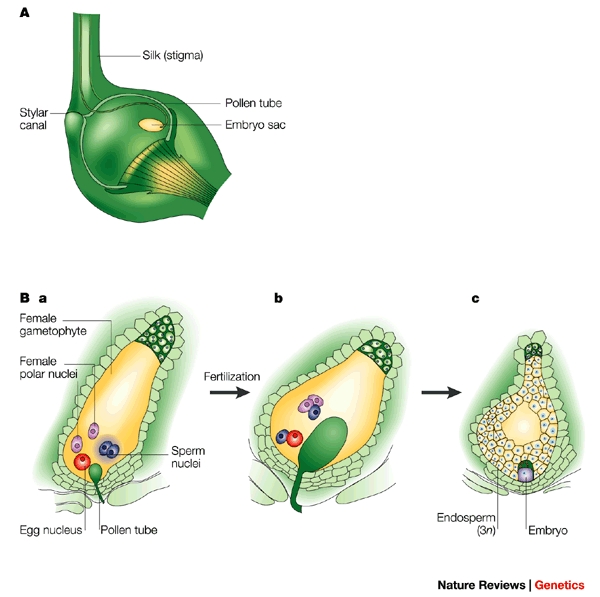
Following fertilization, the flower transforms:
- After fertilization, the ovule develops into a seed.
- After fertilization, the ovary wall thickens and develops into a fruit, which serves to protect the seeds and aid in their dispersal.
Because plants cannot run away from danger or walk toward food, they must alter their growth patterns and chemistry to survive.
3.1 Tropisms: Directional Growth
A tropism is a plant growth response toward or away from a directional stimulus.
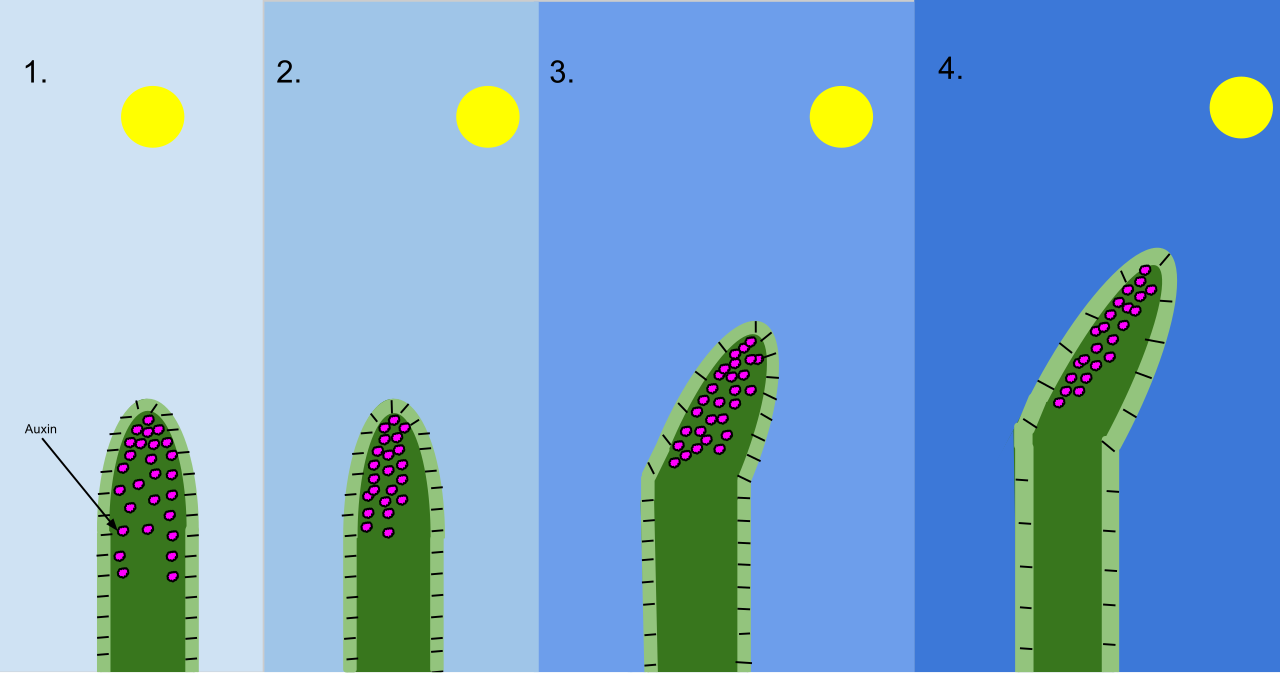
- Phototropism is the directional growth of a plant in response to light.
- Gravitropism is the directional growth of a plant in response to gravity.
- Plant roots exhibit positive gravitropism by growing downward.
- Plant shoots exhibit negative gravitropism by growing upward.
How does a plant "know" which way is down? Statoliths are specialized plastids containing dense starch grains that help plants detect gravity. They settle at the bottom of cells, triggering calcium and hormone signaling that redirects growth.
- Thigmotropism is the directional growth of a plant in response to touch. For example, vines wrapping around a trellis demonstrate positive thigmotropism.
3.2 The Chemical Conductors: Plant Hormones
To execute these movements, plants use chemical messengers. Plant hormones coordinate growth, development, and responses to environmental stimuli. You must know the "Big Five" hormone classes:
| Hormone | Primary Functions & Mechanisms |
|---|---|
| Auxins | Auxins stimulate cell elongation in developing plant shoots. Indoleacetic acid is the most common naturally occurring auxin in plants. <br><br>Auxins mediate phototropism by accumulating on the shaded side of a stem. The accumulation of auxins on the shaded side of a stem causes the stem to bend toward light (because the shaded cells grow longer than the lit cells). <br><br>Additionally, apical dominance is the inhibition of lateral bud growth by auxins produced at the apical meristem, resulting in plants growing taller rather than wider. |
| Cytokinins | Cytokinins are plant hormones that promote cell division. They are the anti-aging hormones of plants and cytokinins act antagonistically to auxins in regulating apical dominance (if you cut off the top of a plant, removing auxin, cytokinins cause the plant to bush out). |
| Gibberellins | Gibberellins stimulate stem elongation, seed germination, and fruit growth. They are the "awaken and grow huge" hormones (frequently sprayed on grocery store grapes to make them larger). |
| Abscisic Acid (ABA) | The stress manager. Abscisic acid slows plant growth and promotes seed dormancy. Crucially, abscisic acid triggers stomatal closure during periods of drought stress to minimize water loss. |
| Ethylene | The only gaseous hormone. Ethylene is a gaseous plant hormone that promotes fruit ripening (why one rotten apple spoils the barrel). <br><br>Ethylene regulates leaf abscission in autumn, allowing trees to drop their leaves safely. <br><br>When a seedling pushes through the soil and hits a rock, the triple response is a growth maneuver induced by ethylene that enables a seedling to avoid physical obstacles (it slows stem elongation, thickens the stem, and causes it to grow horizontally). |
3.3 Fortress Earth: Plant Defenses
Because they are anchored in place, plants are constant targets for herbivores and pathogens. They fight back through multiple layers of defense.
First Line of Defense:
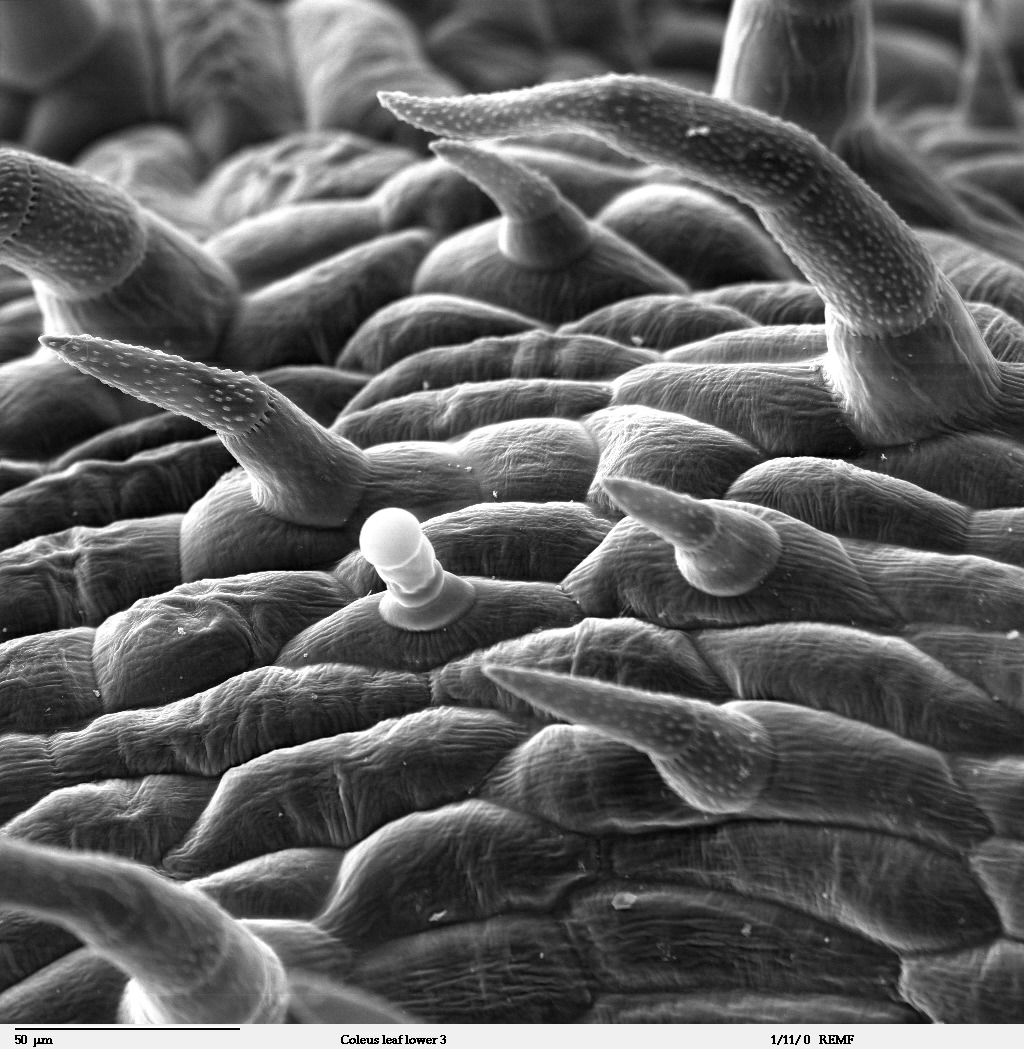
- Plants defend against herbivores using physical barriers such as thorns, spines, and trichomes (tiny, hair-like projections that can trap insects or puncture their exoskeletons).
Second Line of Defense:
- When armor fails, plants use chemical warfare. Plants produce secondary metabolites such as alkaloids and tannins to deter herbivores. Alkaloids (like caffeine and nicotine) are toxic or neurologically disruptive to insects, while tannins bind to digestive enzymes, making the plant violently indigestible.
Anti-Pathogen Defenses: When a virus, bacterium, or fungus breaches the plant, it triggers targeted immune responses:
- The hypersensitive response is a localized defense mechanism that causes cell death at the site of a pathogen infection. The plant deliberately starves the pathogen by killing its own surrounding tissue, effectively burning the bridge to prevent the infection from spreading.

- Before the infected cells die, they send an alarm signal to the rest of the plant. Salicylic acid is a signaling molecule essential for the induction of systemic acquired resistance in plants. (This is the same active compound historically derived from willow bark that we use to make aspirin).
- This signal initiates systemic acquired resistance, [which] provides long-lasting, whole-plant protection against a broad spectrum of pathogens, ensuring the unharmed leaves are chemically fortified before the pathogen can ever reach them.
Teaching Synthesis
When you step in front of your future classroom, remember that biology is not a static list of vocabulary words. The story of plants is a masterclass in evolutionary engineering. By connecting the abstract chemistry of transpiration to the towering heights of redwoods, or the microscopic dance of double fertilization to the fruits in their lunchboxes, you transform rote memorization into a lasting appreciation for the living world.